

Obesity promotes breast epithelium DNA damage in women carrying a germline mutation in BRCA1 or BRCA2
- PMID: 36812344
- PMCID: PMC10557057
- DOI: 10.1126/scitranslmed.ade1857
Obesity promotes breast epithelium DNA damage in women carrying a germline mutation in BRCA1 or BRCA2
Abstract
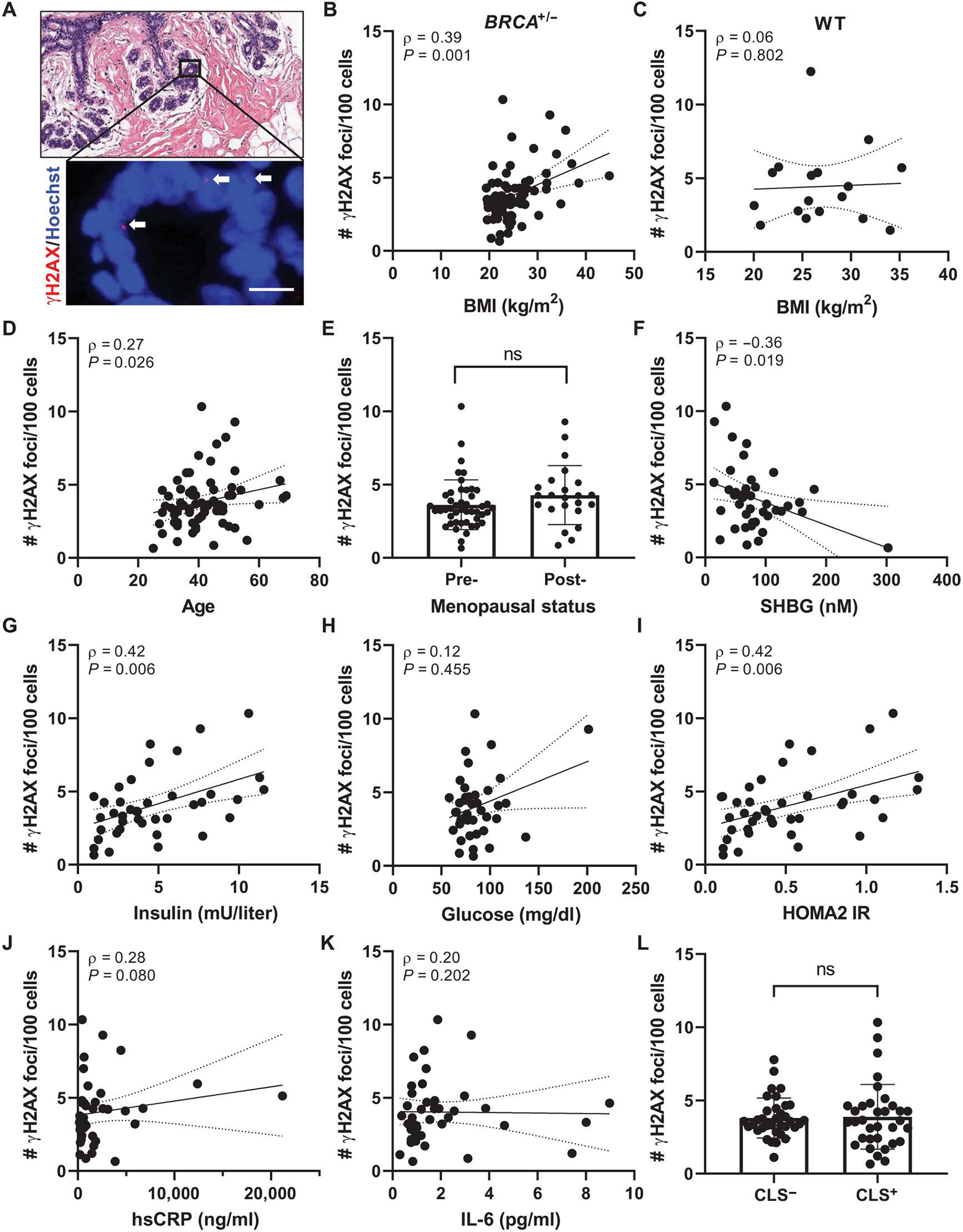
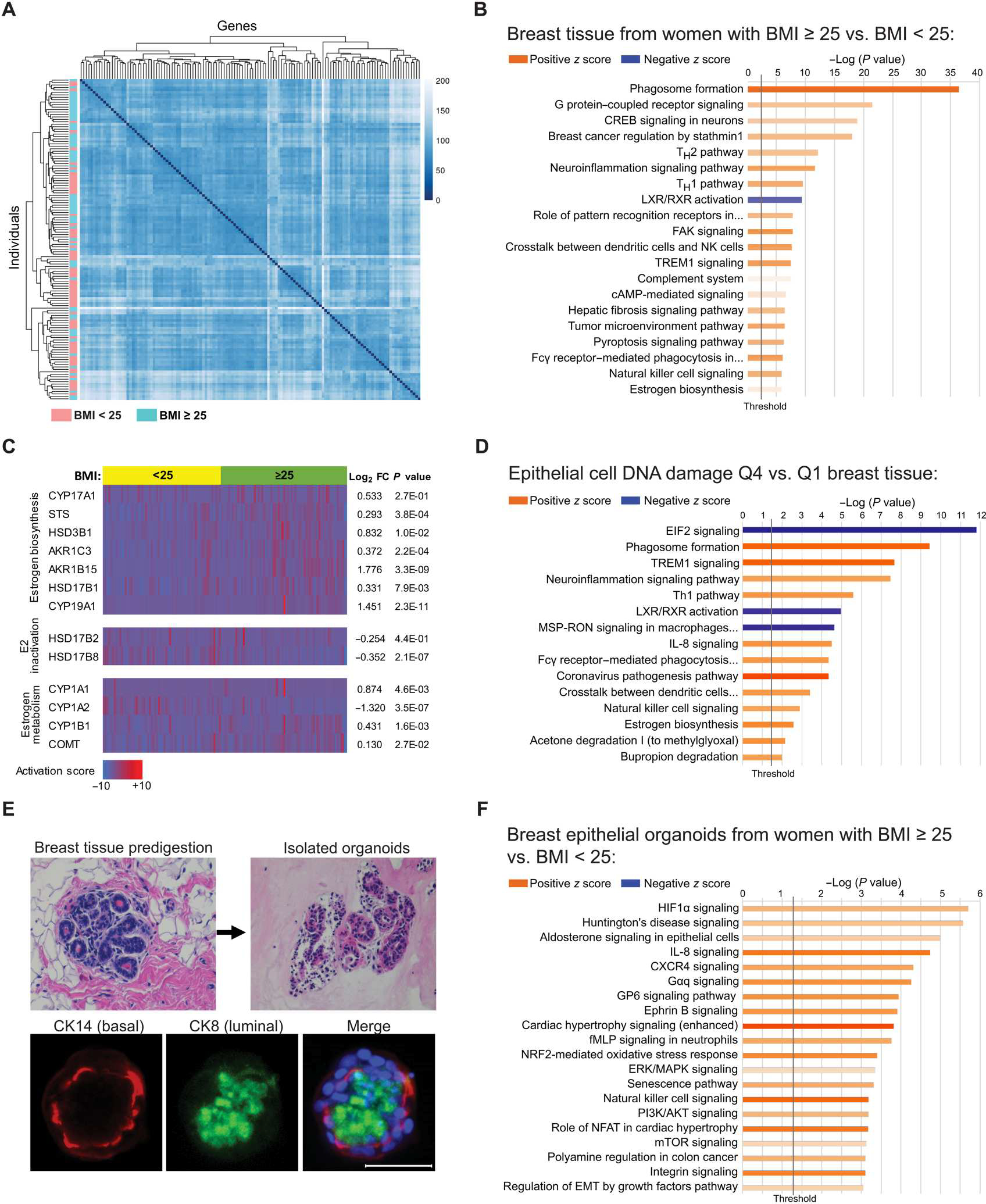
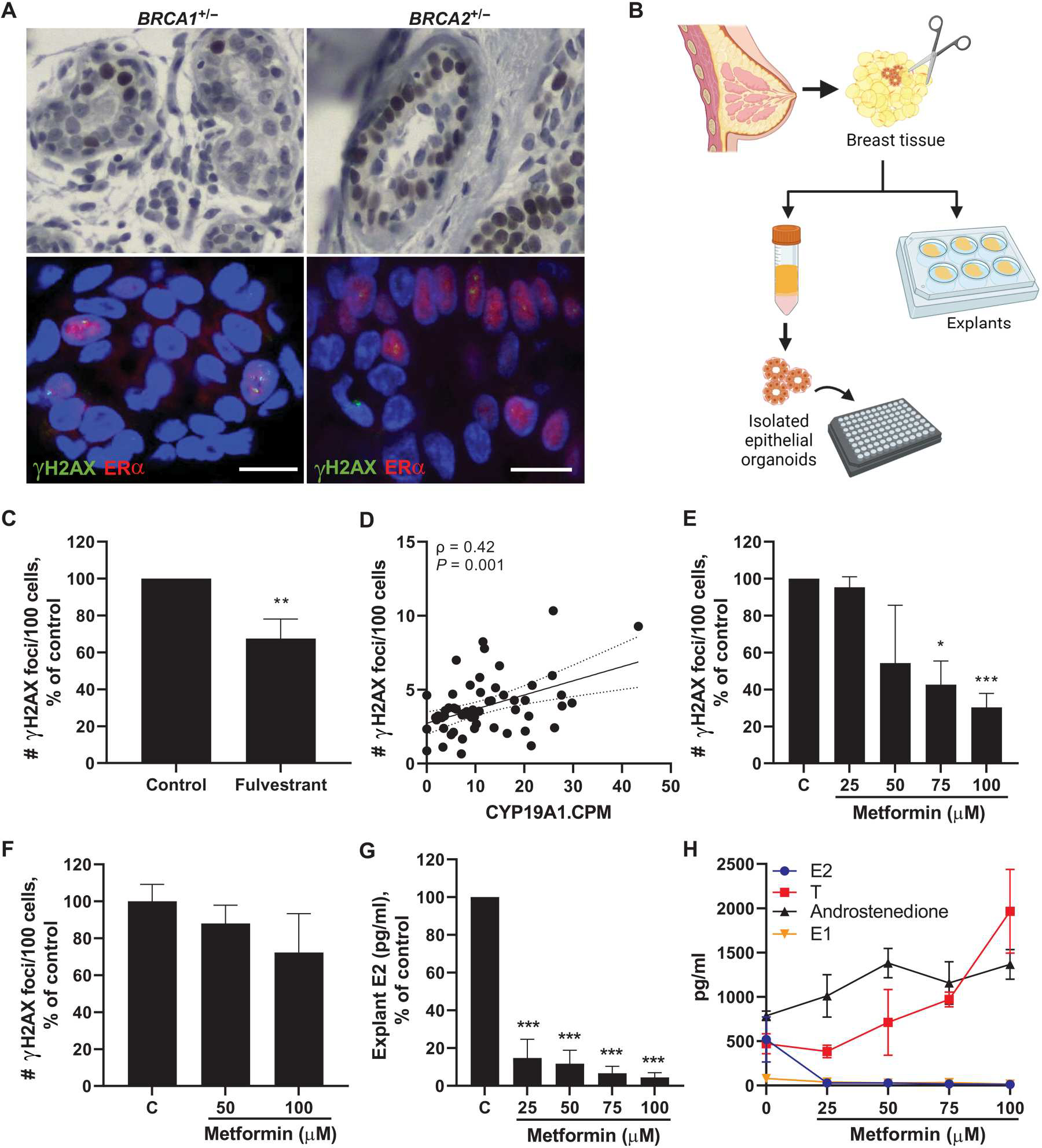
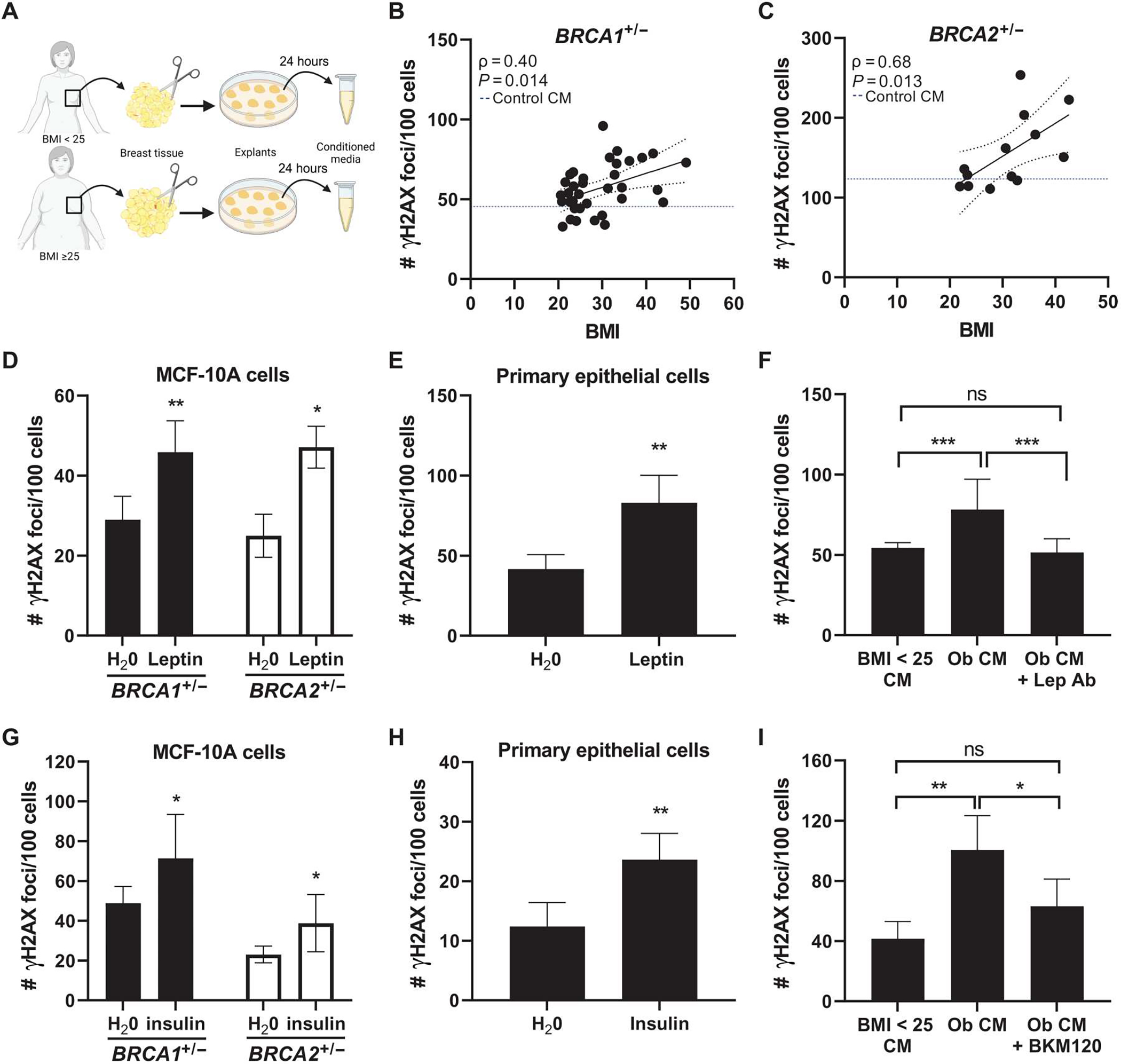
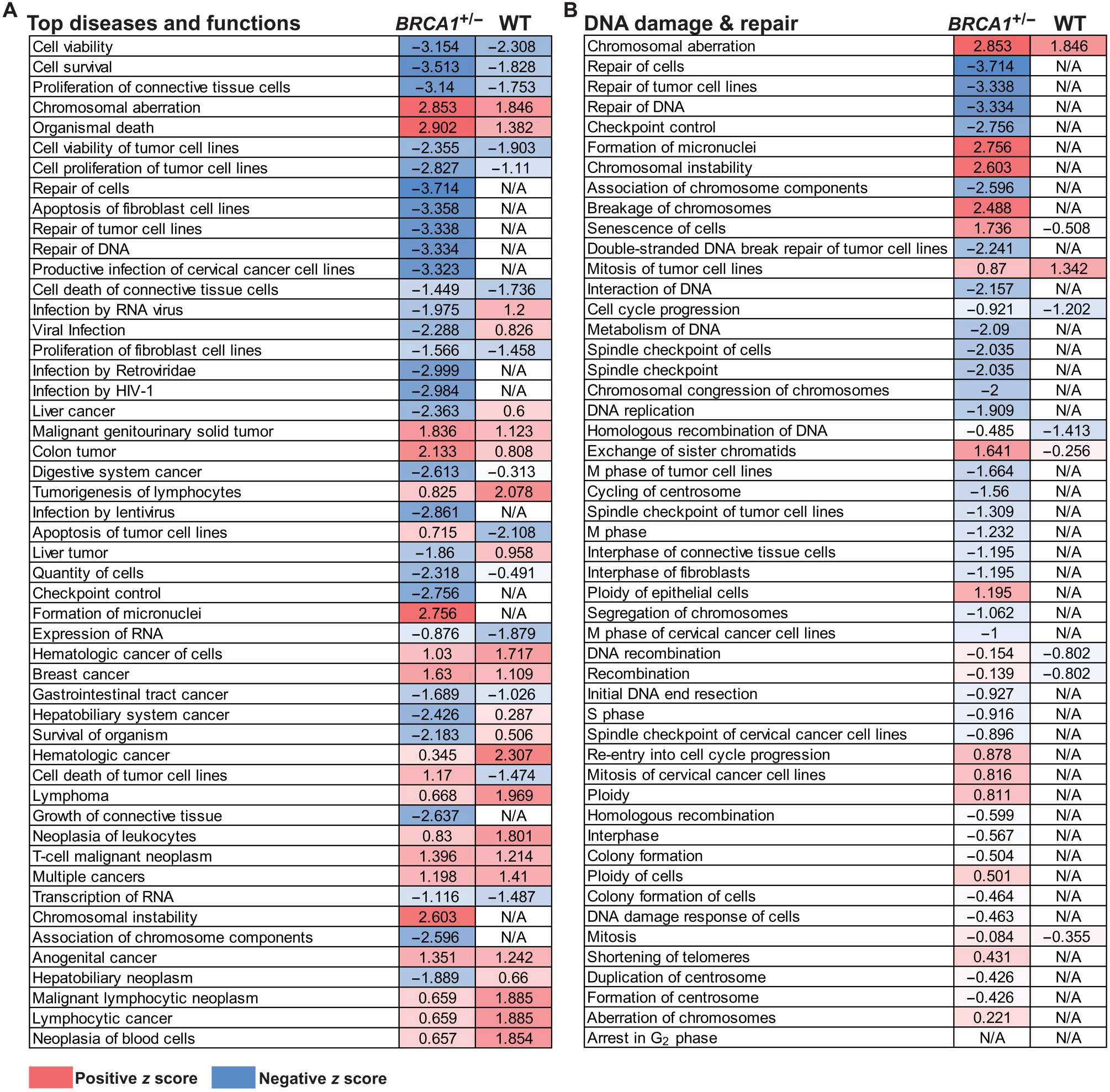
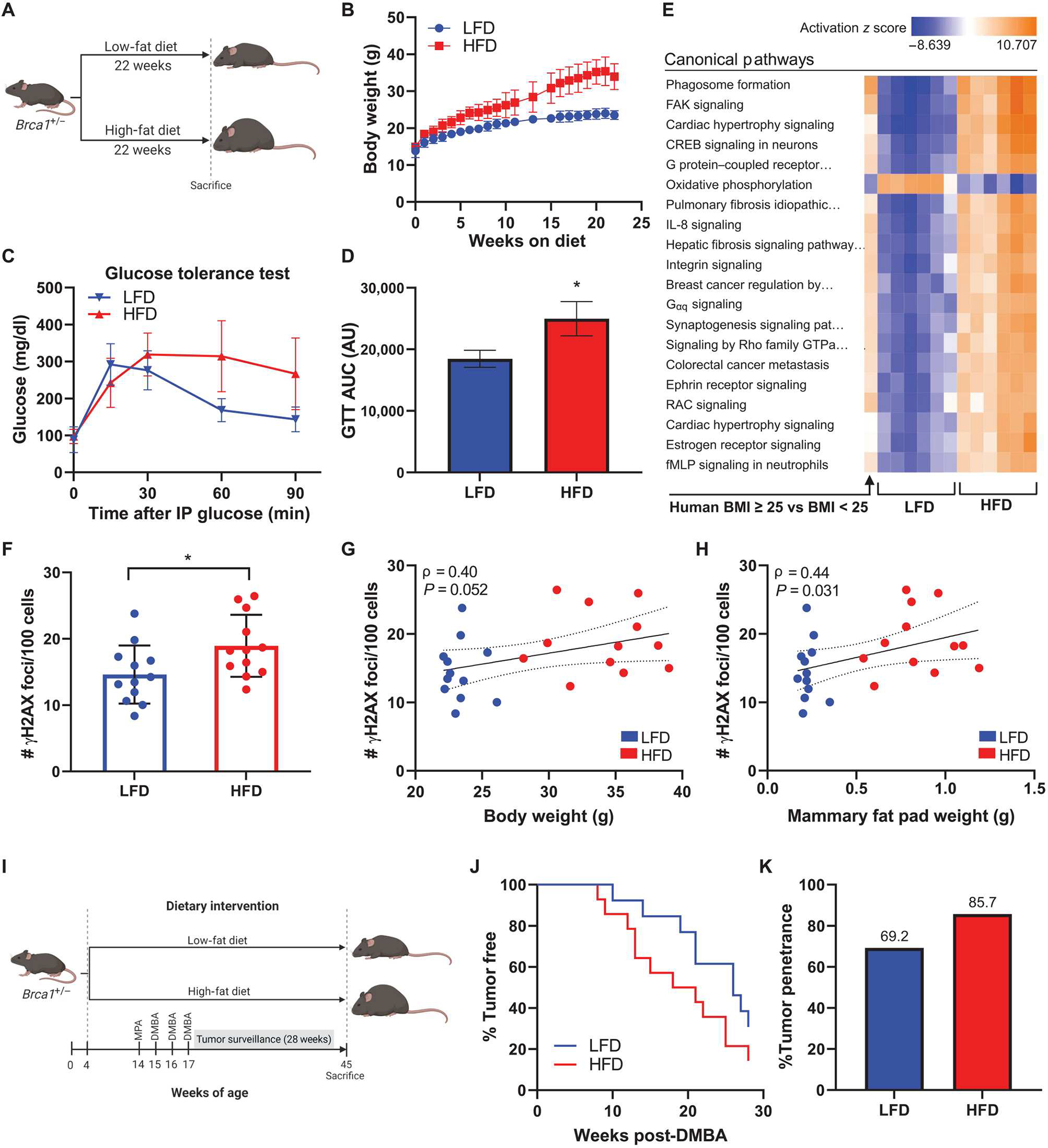
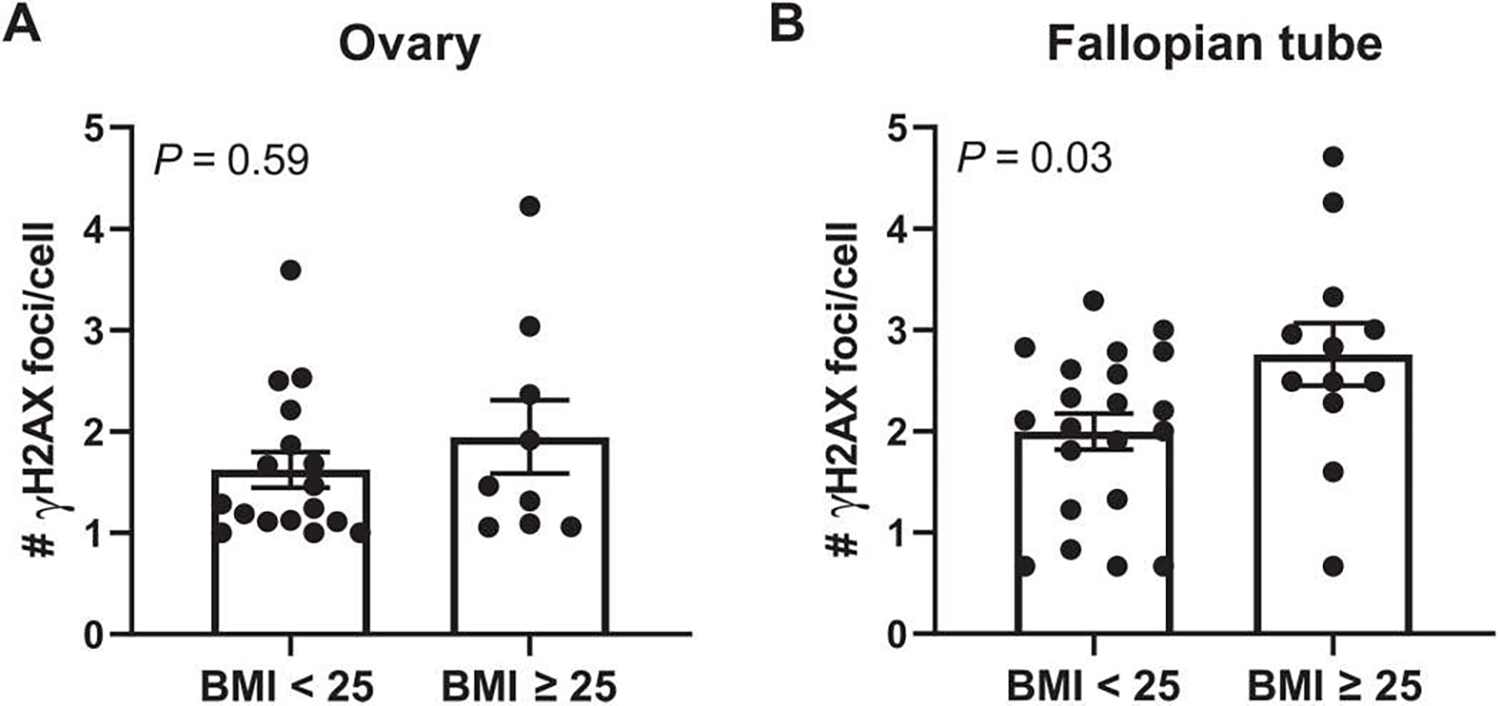
Obesity, defined as a body mass index (BMI) ≥ 30, is an established risk factor for breast cancer among women in the general population after menopause. Whether elevated BMI is a risk factor for women with a germline mutation in BRCA1 or BRCA2 is less clear because of inconsistent findings from epidemiological studies and a lack of mechanistic studies in this population. Here, we show that DNA damage in normal breast epithelia of women carrying a BRCA mutation is positively correlated with BMI and with biomarkers of metabolic dysfunction. In addition, RNA sequencing showed obesity-associated alterations to the breast adipose microenvironment of BRCA mutation carriers, including activation of estrogen biosynthesis, which affected neighboring breast epithelial cells. In breast tissue explants cultured from women carrying a BRCA mutation, we found that blockade of estrogen biosynthesis or estrogen receptor activity decreased DNA damage. Additional obesity-associated factors, including leptin and insulin, increased DNA damage in human BRCA heterozygous epithelial cells, and inhibiting the signaling of these factors with a leptin-neutralizing antibody or PI3K inhibitor, respectively, decreased DNA damage. Furthermore, we show that increased adiposity was associated with mammary gland DNA damage and increased penetrance of mammary tumors in Brca1+/- mice. Overall, our results provide mechanistic evidence in support of a link between elevated BMI and breast cancer development in BRCA mutation carriers. This suggests that maintaining a lower body weight or pharmacologically targeting estrogen or metabolic dysfunction may reduce the risk of breast cancer in this population.
Figures
Comment in
-
Obesity as a promoter in BRCA mutation carriers.Nat Rev Cancer. 2023 May;23(5):272. doi: 10.1038/s41568-023-00564-8. Nat Rev Cancer. 2023. PMID: 36977944 No abstract available.
References
-
- Kuchenbaecker KB, Hopper JL, Barnes DR, Phillips K-A, Mooij TM, Roos-Blom M-J, Jervis S, Van Leeuwen FE, Milne RL, Andrieu N, Goldgar DE, Terry MB, Rookus MA, Easton DF, Antoniou AC, Mcguffog L, Evans DG, Barrowdale D, Frost D, Adlard J, Ong KR, Izatt L, Tischkowitz M, Eeles R, Davidson R, Hodgson S, Ellis S, Nogues C, Lasset C, Stoppa-Lyonnet D, Fricker J-P, Faivre L, Berthet P, Hooning MJ, Van Der Kolk LE, Kets CM, Adank MA, John EM, Chung WK, Andrulis IL, Southey M, Daly MB, Buys SS, Osorio A, Engel C, Kast K, Schmutzler RK, Caldes T, Jakubowska A, Simard J, Friedlander ML, Mclachlan S-A, Machackova E, Foretova L, Tan YY, Singer CF, Olah E, Gerdes A-M, Arver B, Olsson H, Risks of breast, ovarian, and contralateral breast cancer for BRCA1 and BRCA2 mutation carriers. JAMA 317, 2402–2416 (2017). - PubMed
-
- King MC, Marks JH, Mandell JB, New G, Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science 302, 643–646 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
