

The 5-lipoxygenase/cyclooxygenase-2 cross-over metabolite, hemiketal E2, enhances VEGFR2 activation and promotes angiogenesis
- PMID: 36813233
- PMCID: PMC10040730
- DOI: 10.1016/j.jbc.2023.103050
The 5-lipoxygenase/cyclooxygenase-2 cross-over metabolite, hemiketal E2, enhances VEGFR2 activation and promotes angiogenesis
Abstract
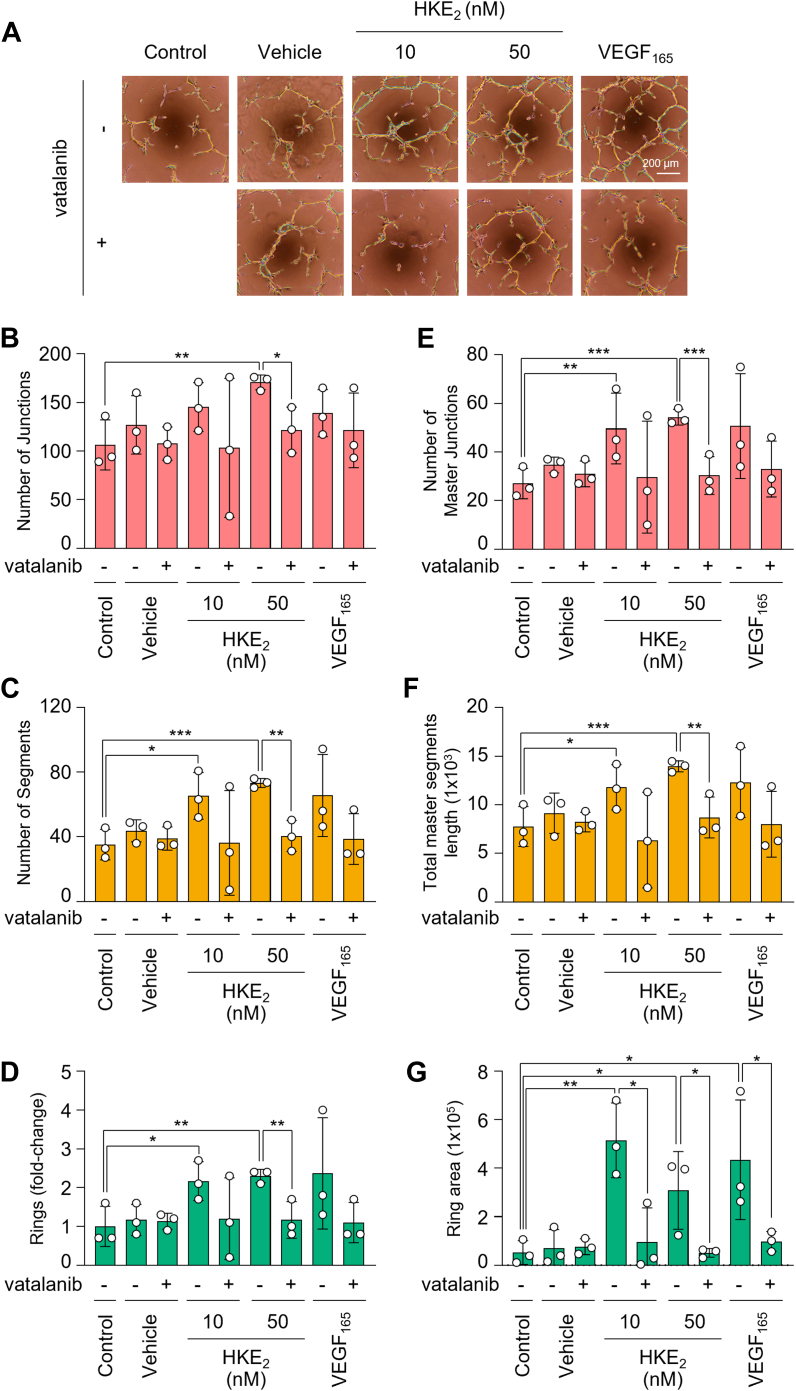
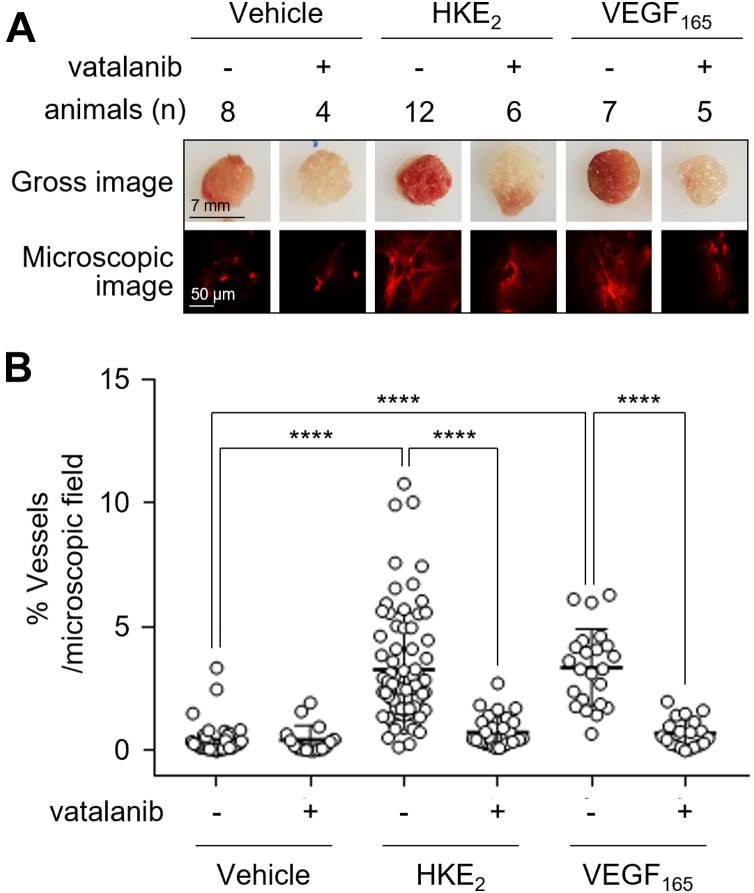
Consecutive oxygenation of arachidonic acid by 5-lipoxygenase and cyclooxygenase-2 yields the hemiketal eicosanoids, HKE2 and HKD2. Hemiketals stimulate angiogenesis by inducing endothelial cell tubulogenesis in culture; however, how this process is regulated has not been determined. Here, we identify vascular endothelial growth factor receptor 2 (VEGFR2) as a mediator of HKE2-induced angiogenesis in vitro and in vivo. We found that HKE2 treatment of human umbilical vein endothelial cells dose-dependently increased the phosphorylation of VEGFR2 and the downstream kinases ERK and Akt that mediated endothelial cell tubulogenesis. In vivo, HKE2 induced the growth of blood vessels into polyacetal sponges implanted in mice. HKE2-mediated effects in vitro and in vivo were blocked by the VEGFR2 inhibitor vatalanib, indicating that the pro-angiogenic effect of HKE2 was mediated by VEGFR2. HKE2 covalently bound and inhibited PTP1B, a protein tyrosine phosphatase that dephosphorylates VEGFR2, thereby providing a possible molecular mechanism for how HKE2 induced pro-angiogenic signaling. In summary, our studies indicate that biosynthetic cross-over of the 5-lipoxygenase and cyclooxygenase-2 pathways gives rise to a potent lipid autacoid that regulates endothelial cell function in vitro and in vivo. These findings suggest that common drugs targeting the arachidonic acid pathway could prove useful in antiangiogenic therapy.
Keywords: 5-lipoxygenase; arachidonic acid; cyclooxygenase-2; eicosanoid; endothelial cell; kinase signaling; sponge assay.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
