

NOX2 inhibition enables retention of the circadian clock in BV2 microglia and primary macrophages
- PMID: 36814920
- PMCID: PMC9939898
- DOI: 10.3389/fimmu.2023.1106515
NOX2 inhibition enables retention of the circadian clock in BV2 microglia and primary macrophages
Abstract
Introduction: Sustained neuroinflammation is a major contributor to the progression of neurodegenerative diseases such as Alzheimer's (AD) and Parkinson's (PD) diseases. Neuroinflammation, like other cellular processes, is affected by the circadian clock. Microglia, the resident immune cells in the brain, act as major contributors to neuroinflammation and are under the influence of the circadian clock. Microglial responses such as activation, recruitment, and cytokine expression are rhythmic in their response to various stimuli. While the link between circadian rhythms and neuroinflammation is clear, significant gaps remain in our understanding of this complex relationship. To gain a greater understanding of this relationship, the interaction between the microglial circadian clock and the enzyme NADPH Oxidase Isoform 2 (NOX2) was studied; NOX2 is essential for the production of reactive oxygen species (ROS) in oxidative stress, an integral characteristic of neuroinflammation.
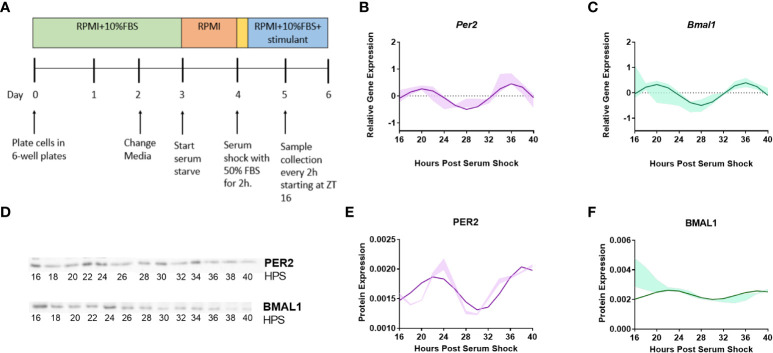
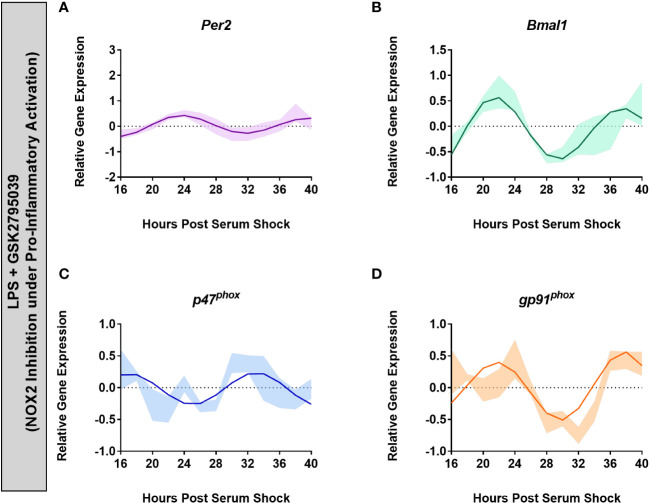
Methods: BV2 microglia were examined over circadian time, demonstrating oscillations of the clock genes Per2 and Bmal1 and the NOX2 subunits gp91phox and p47phox.
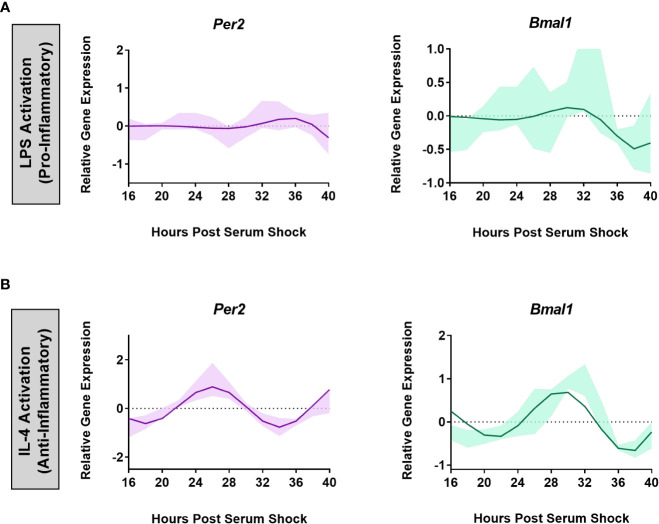
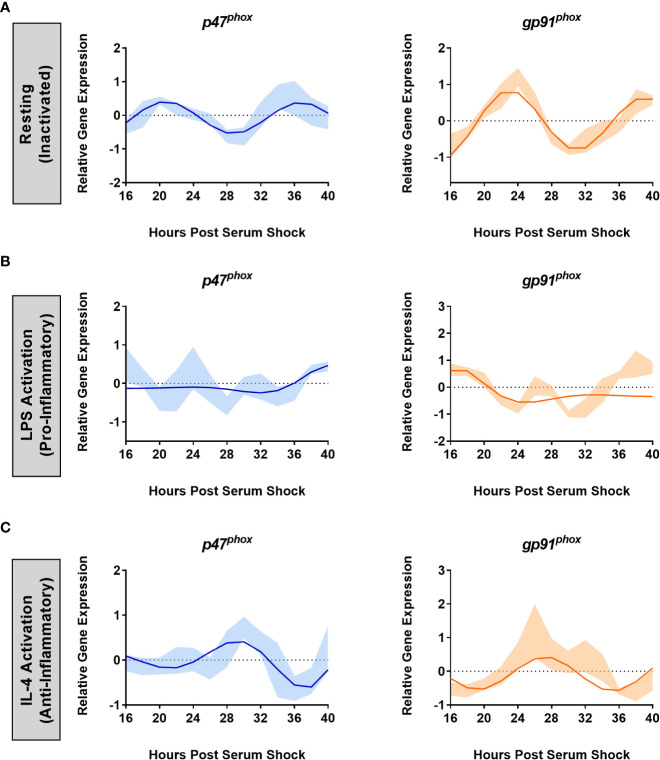
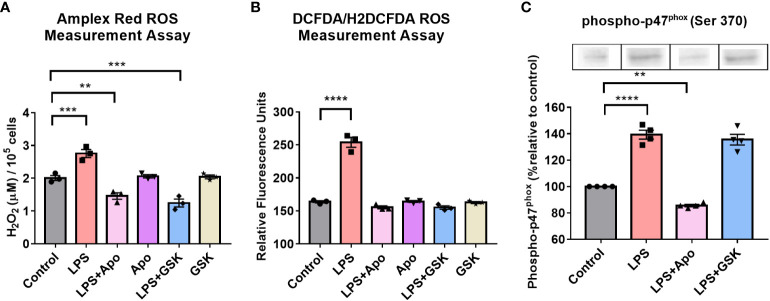
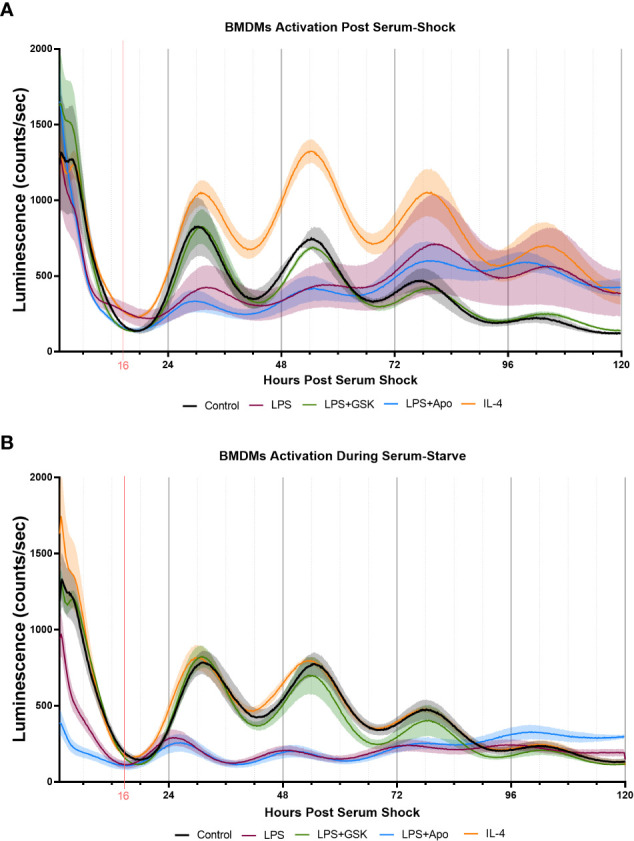
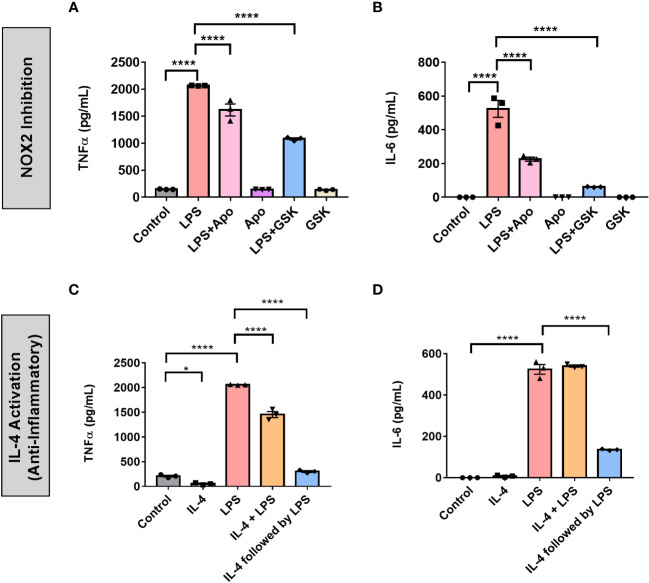
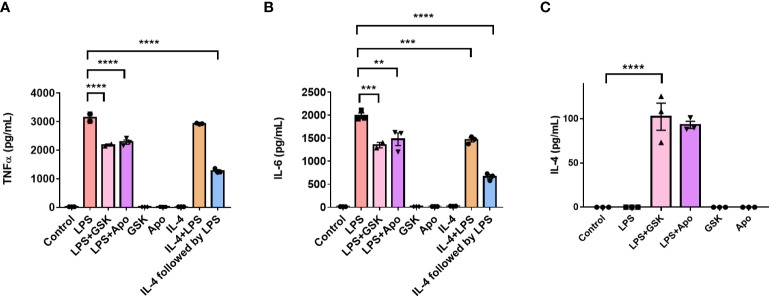
Results: The BV2 microglial clock exerted significant control over NOX2 expression and inhibition of NOX2 enabled the microglia to retain a functional circadian clock while reducing levels of ROS and inflammatory cytokines. These trends were mirrored in mouse bone marrow-derived primary macrophages.
Conclusions: NOX2 plays a crucial role in the interaction between the circadian clock and the activation of microglia/macrophages into their pro-inflammatory state, which has important implications in the control of neuroinflammation.
Keywords: BV2 microglia; NOX2; circadian rhythm; neuroinflammation; oxidative stress; primary macrophages.
Copyright © 2023 Muthukumarasamy, Buel, Hurley and Dordick.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The circadian clock protein Rev-erbα provides neuroprotection and attenuates neuroinflammation against Parkinson's disease via the microglial NLRP3 inflammasome.J Neuroinflammation. 2022 Jun 6;19(1):133. doi: 10.1186/s12974-022-02494-y. J Neuroinflammation. 2022. PMID: 35668454 Free PMC article.
-
Microglial clock dysfunction during neuroinflammation impairs oligodendrocyte progenitor cell recruitment and disrupts neuroimmune homeostasis.Front Immunol. 2025 Jul 7;16:1620343. doi: 10.3389/fimmu.2025.1620343. eCollection 2025. Front Immunol. 2025. PMID: 40692797 Free PMC article.
-
Complement C3 Enhances LPS-Elicited Neuroinflammation and Neurodegeneration Via the Mac1/NOX2 Pathway.Mol Neurobiol. 2023 Sep;60(9):5167-5183. doi: 10.1007/s12035-023-03393-w. Epub 2023 Jun 3. Mol Neurobiol. 2023. PMID: 37268807 Free PMC article.
-
Circadian rhythm, microglia-mediated neuroinflammation, and Alzheimer's disease.Neurosci Biobehav Rev. 2025 Mar;170:106044. doi: 10.1016/j.neubiorev.2025.106044. Epub 2025 Feb 4. Neurosci Biobehav Rev. 2025. PMID: 39914702 Review.
-
Circadian regulation of microglia function: Potential targets for treatment of Parkinson's Disease.Ageing Res Rev. 2024 Mar;95:102232. doi: 10.1016/j.arr.2024.102232. Epub 2024 Feb 15. Ageing Res Rev. 2024. PMID: 38364915 Review.
Cited by
-
Neuronal and glial vulnerability of the suprachiasmatic nucleus in tauopathies: evidence from human studies and animal models.Mol Neurodegener. 2024 Jan 10;19(1):4. doi: 10.1186/s13024-023-00695-4. Mol Neurodegener. 2024. PMID: 38195580 Free PMC article. Review.
-
Involvement of circadian clock protein PER2 in controlling sleep deprivation induced HMGB1 up-regulation by targeting p300 in the cortex.Sci Rep. 2025 Apr 10;15(1):12253. doi: 10.1038/s41598-025-96931-6. Sci Rep. 2025. PMID: 40210902 Free PMC article.
-
Circadian rhythm disruption: a potential trigger in Parkinson's disease pathogenesis.Front Cell Neurosci. 2024 Oct 30;18:1464595. doi: 10.3389/fncel.2024.1464595. eCollection 2024. Front Cell Neurosci. 2024. PMID: 39539340 Free PMC article. Review.
-
New insight on microglia activation in neurodegenerative diseases and therapeutics.Front Neurosci. 2023 Dec 22;17:1308345. doi: 10.3389/fnins.2023.1308345. eCollection 2023. Front Neurosci. 2023. PMID: 38188026 Free PMC article. Review.
-
Phosphatidylcholine suppresses inflammatory responses in LPS-stimulated MG6 microglial cells by inhibiting NF-κB/JNK/p38 MAPK signaling.PLoS One. 2025 Jul 28;20(7):e0328206. doi: 10.1371/journal.pone.0328206. eCollection 2025. PLoS One. 2025. PMID: 40720378 Free PMC article.
References
-
- Kigerl KA, Gensel JC, Ankeny DP, Alexander JK, Donnelly DJ, Popovich PG. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J Neurosci (2009) 29(43):13435–44. doi: 10.1523/JNEUROSCI.3257-09.2009 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous