

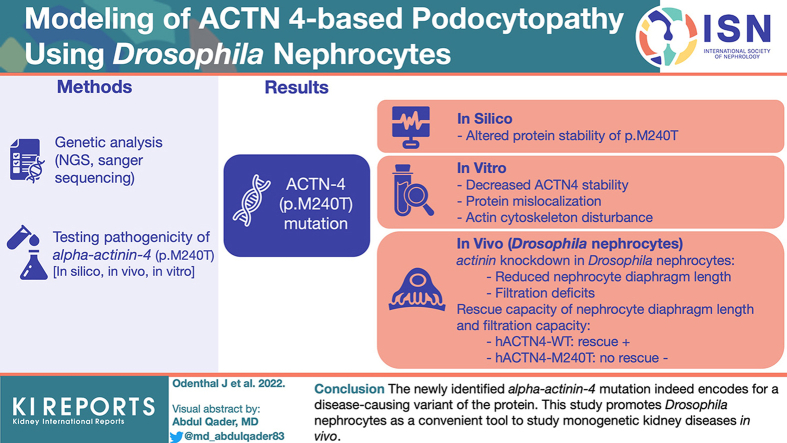
Modeling of ACTN4-Based Podocytopathy Using Drosophila Nephrocytes
- PMID: 36815115
- PMCID: PMC9939316
- DOI: 10.1016/j.ekir.2022.10.024
Modeling of ACTN4-Based Podocytopathy Using Drosophila Nephrocytes
Abstract
Introduction: Genetic disorders are among the most prevalent causes leading to progressive glomerular disease and, ultimately, end-stage renal disease (ESRD) in children and adolescents. Identification of underlying genetic causes is indispensable for targeted treatment strategies and counseling of affected patients and their families.
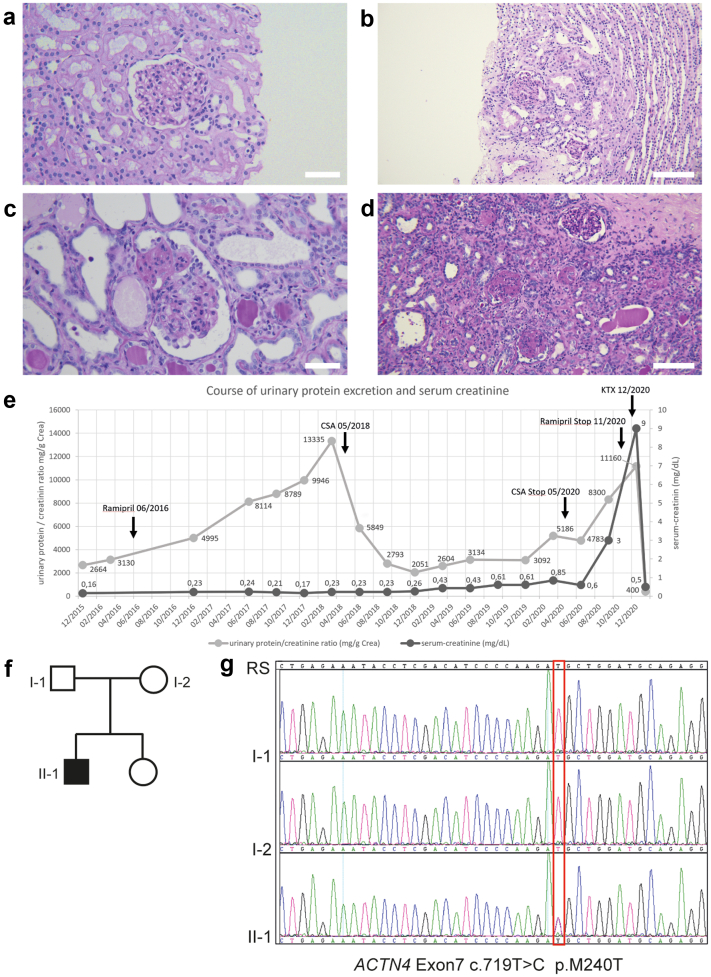
Methods: Here, we report on a boy who presented at 4 years of age with proteinuria and biopsy-proven focal segmental glomerulosclerosis (FSGS) that was temporarily responsive to treatment with ciclosporin A. Molecular genetic testing identified a novel mutation in alpha-actinin-4 (p.M240T). We describe a feasible and efficient experimental approach to test its pathogenicity by combining in silico, in vitro, and in vivo analyses.
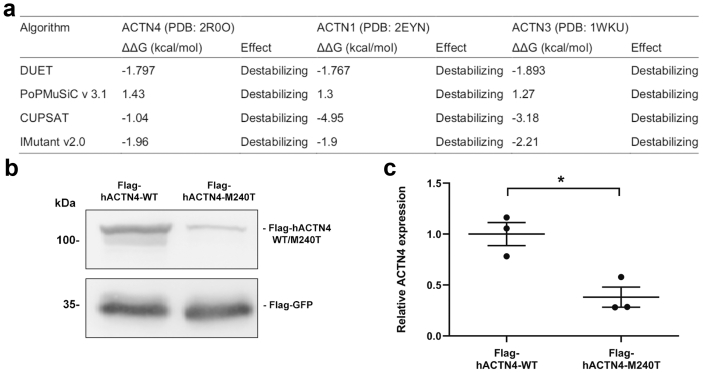
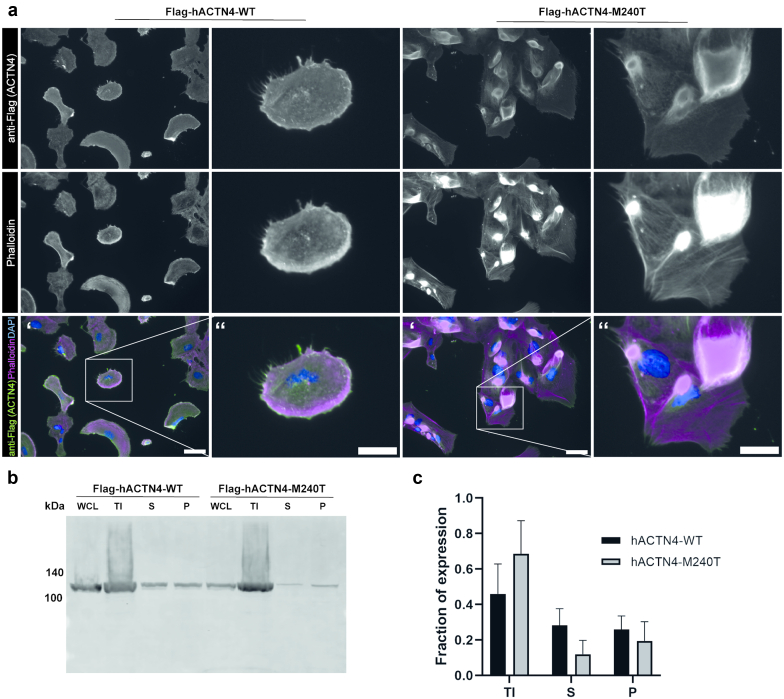
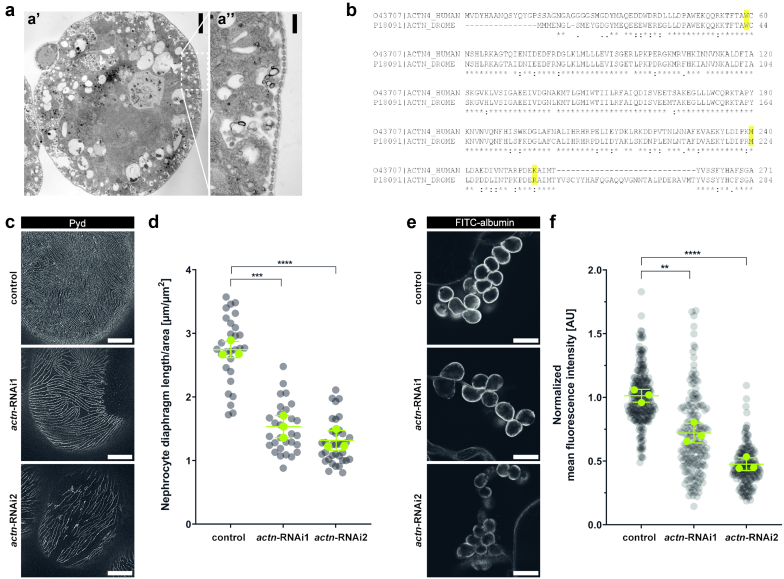
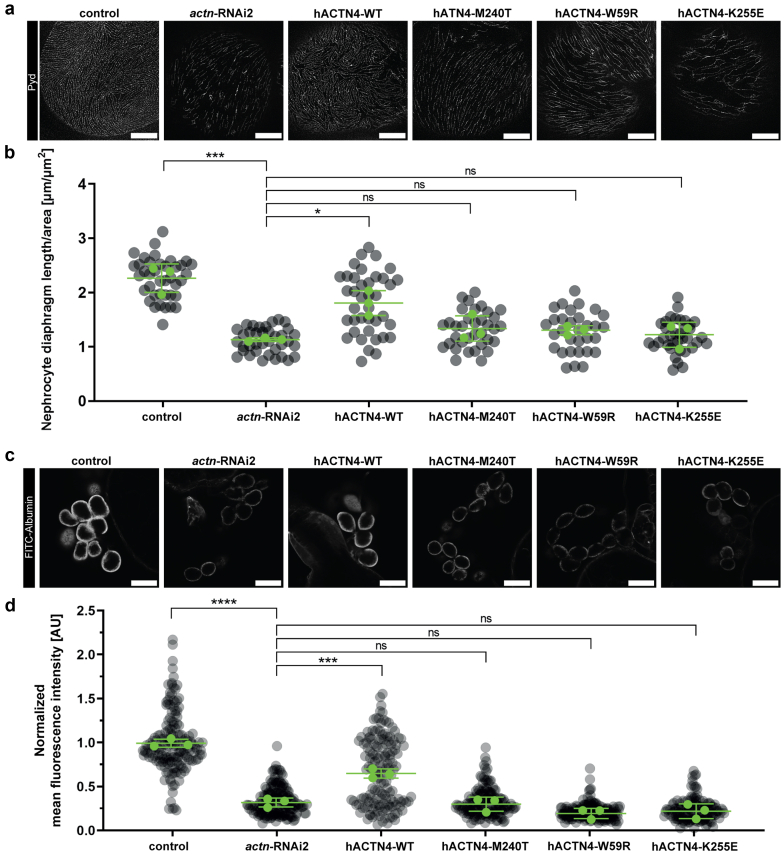
Results: The de novo p.M240T mutation led to decreased alpha-actinin-4 stability as well as protein mislocalization and actin cytoskeleton rearrangements. Transgenic expression of wild-type human alpha-actinin-4 in Drosophila melanogaster nephrocytes was able to ameliorate phenotypes associated with the knockdown of endogenous actinin. In contrast, p.M240T, as well as other established disease variants p.W59R and p.K255E, failed to rescue these phenotypes, underlining the pathogenicity of the novel alpha-actinin-4 variant.
Conclusion: Our data highlight that the newly identified alpha-actinin-4 mutation indeed encodes for a disease-causing variant of the protein and promote the Drosophila model as a simple and convenient tool to study monogenic kidney disease in vivo.
Keywords: ACTN4; Drosophila; FSGS; nephrocyte; nephrotic syndrome; podocyte.
© 2022 Published by Elsevier Inc. on behalf of the International Society of Nephrology.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
