

Hsp70.1 carbonylation induces lysosomal cell death for lifestyle-related diseases
- PMID: 36819480
- PMCID: PMC9936620
- DOI: 10.3389/fmolb.2022.1063632
Hsp70.1 carbonylation induces lysosomal cell death for lifestyle-related diseases
Abstract
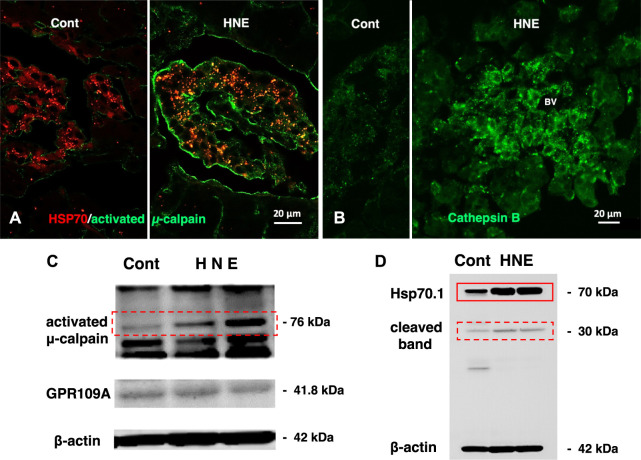
Alzheimer's disease, type 2 diabetes, and non-alcoholic steatohepatitis (NASH) constitute increasingly prevalent disorders. Individuals with type 2 diabetes are well-known to be susceptible to Alzheimer's disease. Although the pathogenesis of each disorder is multifactorial and the causal relation remains poorly understood, reactive oxygen species (ROS)-induced lipid and protein oxidation conceivably plays a common role. Lipid peroxidation product was recently reported to be a key factor also for non-alcoholic steatohepatitis, because of inducing hepatocyte degeneration/death. Here, we focus on implication of the representative lipid-peroxidation product 'hydroxynonenal' for the cell degeneration/death of brain, pancreas, and liver. Since Hsp70.1 has dual roles as a chaperone and lysosomal membrane stabilizer, hydroxynonenal-mediated oxidative injury (carbonylation) of Hsp70.1 was highlighted. After intake of high-fat diets, oxidation of free fatty acids in mitochondria generates ROS which enhance oxidation of ω-6 polyunsaturated fatty acids (PUFA) involved within biomembranes and generate hydroxynonenal. In addition, hydroxynonenal is generated during cooking deep-fried foods with vegetable oils especially containing linoleic acids. These intrinsic and exogenous hydroxynonenal synergically causes an increase in its serum and organ levels to induce Hsp70.1 oxidation. As it is amphiphilic; being water-soluble but displays strong lipophilic characteristics, hydroxynonenal can diffuse within the cells and react with targets like senile and/or atheromatous plaques outside the cells. Hydroxynonenal can deepen and expand lysosomal injuries by facilitating 'calpain-mediated cleavage of the carbonylated Hsp70.1'. Despite the unique anatomical, physiological, and biochemical characteristics of each organ for its specific disease, there should be a common cascade of the cell degeneration/death which is caused by hydroxynonenal. This review aims to implicate hydroxynonenal-mediated Hsp70.1 carbonylation for lysosomal membrane permeabilization/rupture and the resultant cathepsin leakage for inducing cell degeneration/death. Given the tremendous number of worldwide people suffering various lifestyle-related diseases, it is valuable to consider how ω-6 PUFA-rich vegetable oils is implicated for the organ disorder.
Keywords: Alzheimer’s disease; calpain-cathepsin hypothesis; hydroxynonenal; non-alcoholic steatohepatitis; type 2 diabetes.
Copyright © 2023 Yamashima, Seike, Oikawa, Kobayashi, Kido, Yanagi, Yamamiya, Li, Boontem and Mizukoshi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures









References
Publication types
LinkOut - more resources
Full Text Sources
Research Materials