

This is a preprint.
Temporal Notch signaling regulates mucociliary cell fates through Hes-mediated competitive de-repression
- PMID: 36824900
- PMCID: PMC9949065
- DOI: 10.1101/2023.02.15.528675
Temporal Notch signaling regulates mucociliary cell fates through Hes-mediated competitive de-repression
Abstract
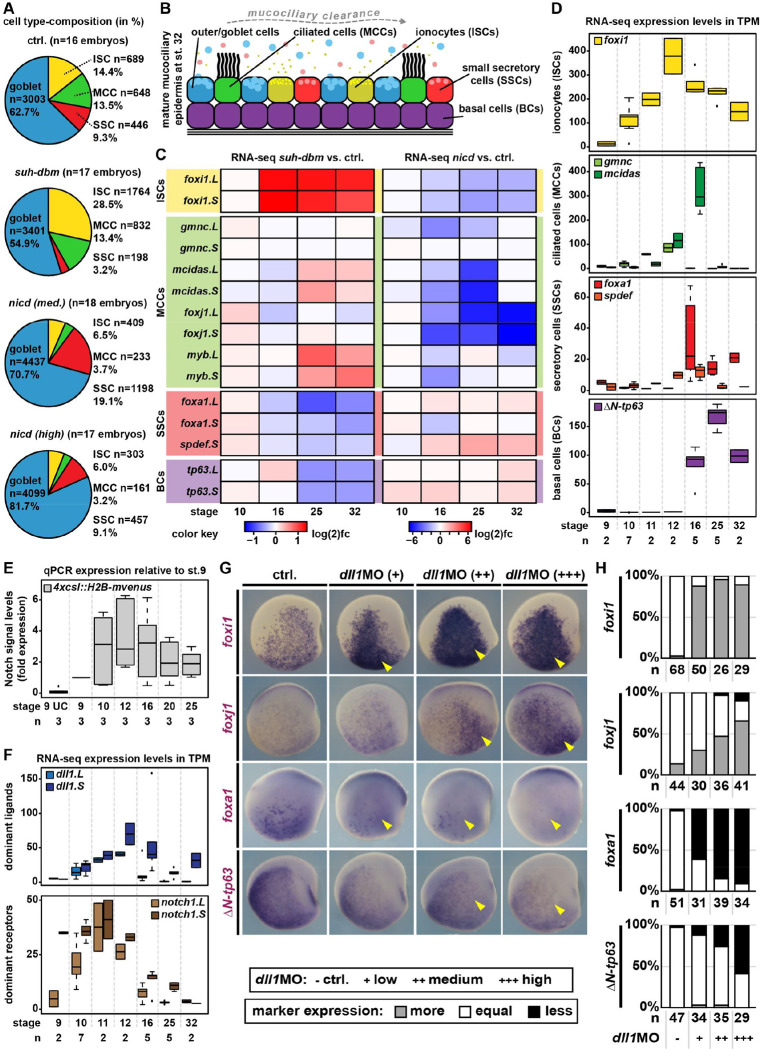
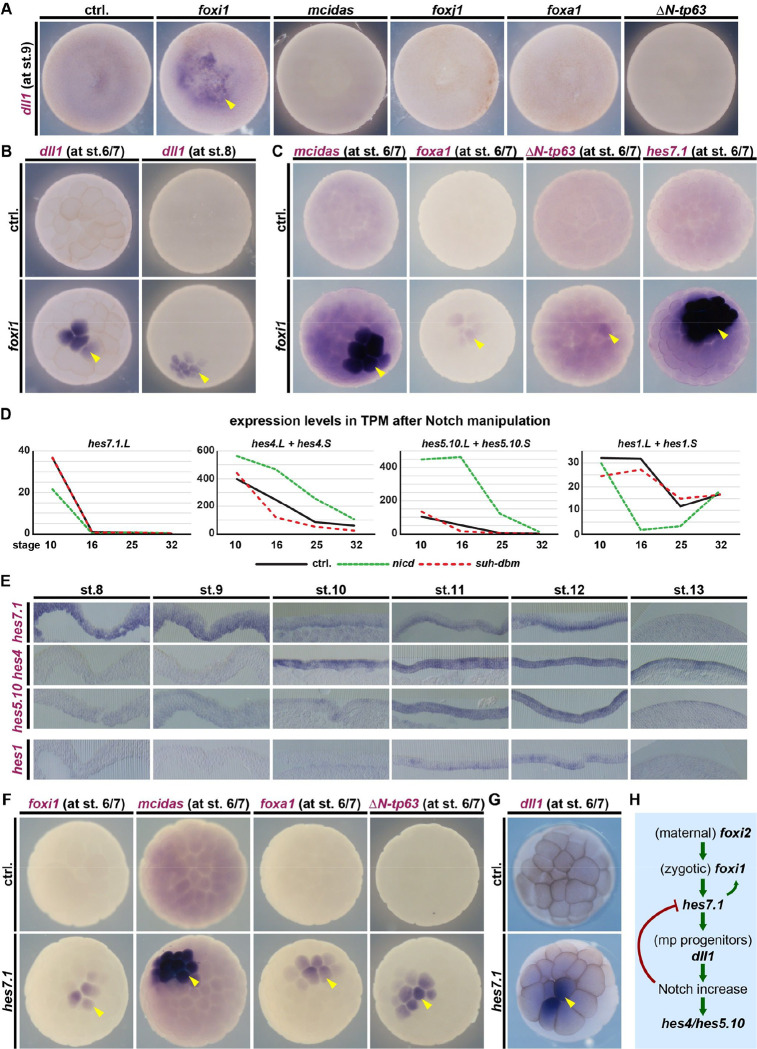
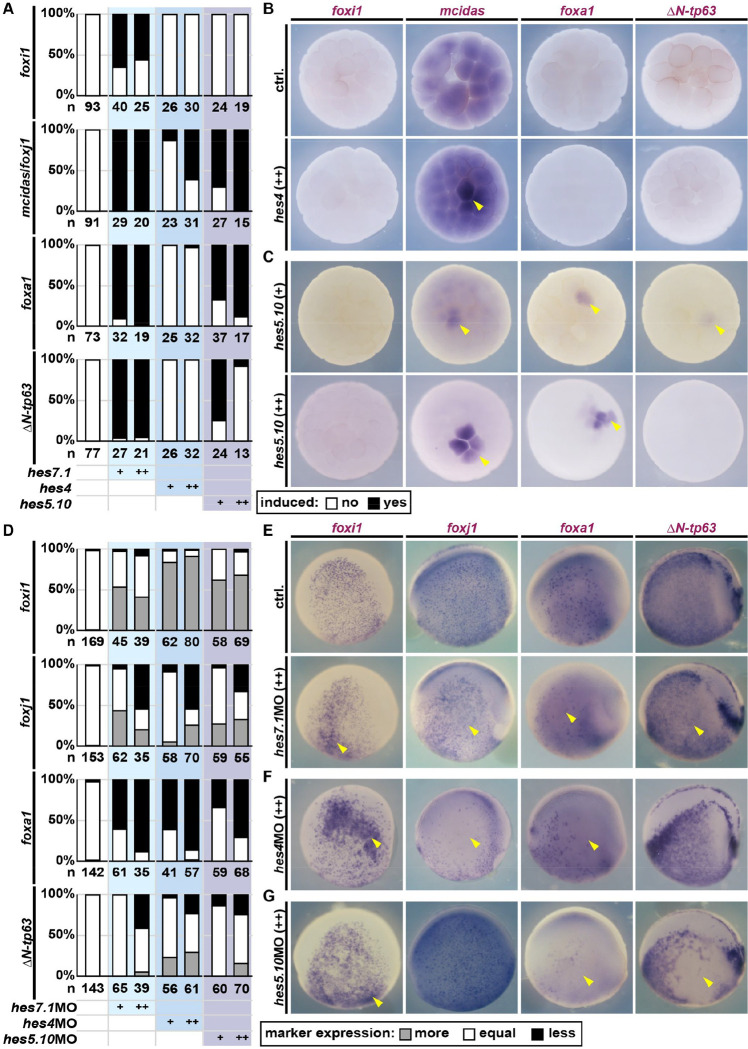
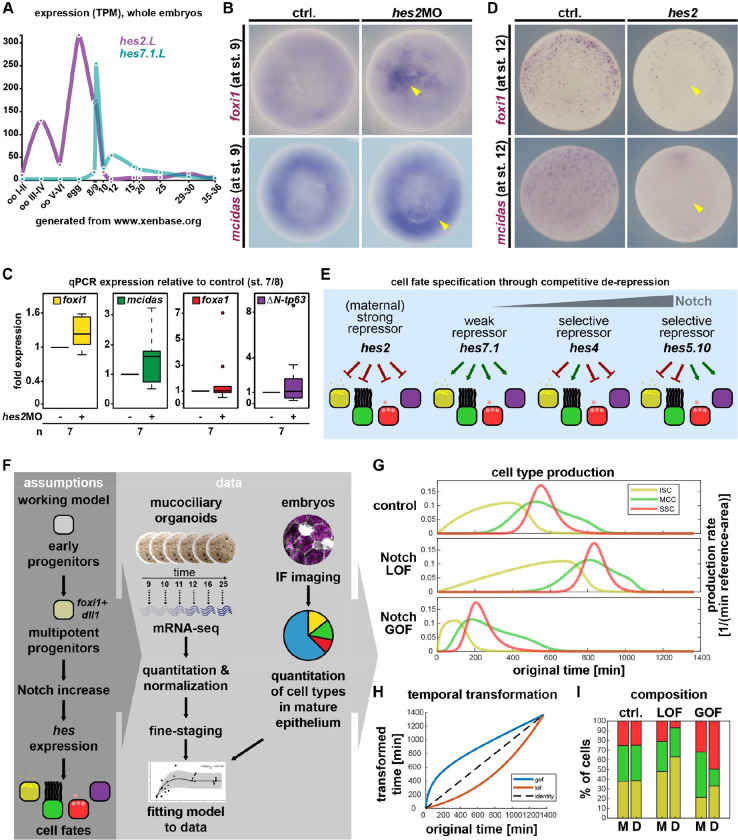
Tissue functions are determined by the types and ratios of cells present, but little is known about self-organizing principles establishing correct cell type compositions. Mucociliary airway clearance relies on the correct balance between secretory and ciliated cells, which is regulated by Notch signaling across mucociliary systems. Using the airway-like Xenopus epidermis, we investigate how cell fates depend on signaling, how signaling levels are controlled, and how Hes transcription factors regulate cell fates. We show that four mucociliary cell types each require different Notch levels and that their specification is initiated sequentially by a temporal Notch gradient. We describe a novel role for Foxi1 in the generation of Delta-expressing multipotent progenitors through Hes7.1. Hes7.1 is a weak repressor of mucociliary genes and overcomes maternal repression by the strong repressor Hes2 to initiate mucociliary development. Increasing Notch signaling then inhibits Hes7.1 and activates first Hes4, then Hes5.10, which selectively repress cell fates. We have uncovered a self-organizing mechanism of mucociliary cell type composition by competitive de-repression of cell fates by a set of differentially acting repressors. Furthermore, we present an in silico model of this process with predictive abilities.
Keywords: Xenopus; airway epithelium; basal cells; development; mathematical modeling; mucus; multi-ciliated cells.
Figures
Similar articles
-
Foxi1 regulates multiple steps of mucociliary development and ionocyte specification through transcriptional and epigenetic mechanisms.bioRxiv [Preprint]. 2024 Nov 4:2024.10.27.620464. doi: 10.1101/2024.10.27.620464. bioRxiv. 2024. PMID: 39484493 Free PMC article. Preprint.
-
Signaling Control of Mucociliary Epithelia: Stem Cells, Cell Fates, and the Plasticity of Cell Identity in Development and Disease.Cells Tissues Organs. 2022;211(6):736-753. doi: 10.1159/000514579. Epub 2021 Apr 26. Cells Tissues Organs. 2022. PMID: 33902038 Free PMC article. Review.
-
Integrative genomic analyses on HES/HEY family: Notch-independent HES1, HES3 transcription in undifferentiated ES cells, and Notch-dependent HES1, HES5, HEY1, HEY2, HEYL transcription in fetal tissues, adult tissues, or cancer.Int J Oncol. 2007 Aug;31(2):461-6. Int J Oncol. 2007. PMID: 17611704
-
Identification and characterization of human HES2, HES3, and HES5 genes in silico.Int J Oncol. 2004 Aug;25(2):529-34. Int J Oncol. 2004. PMID: 15254753
-
Xenopus epidermal and endodermal epithelia as models for mucociliary epithelial evolution, disease, and metaplasia.Genesis. 2021 Feb;59(1-2):e23406. doi: 10.1002/dvg.23406. Epub 2021 Jan 5. Genesis. 2021. PMID: 33400364 Review.
References
-
- Deblandre G. A., Wettstein D. A., Koyano-Nakagawa N. and Kintner C. (1999). A two-step mechanism generates the spacing pattern of the ciliated cells in the skin of Xenopus embryos. Development 126, 4715–4728. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources