

This is a preprint.
Context-dependent activation of a social behavior brain network associates with learned vocal production
- PMID: 36824963
- PMCID: PMC9949236
- DOI: 10.21203/rs.3.rs-2587773/v1
Context-dependent activation of a social behavior brain network associates with learned vocal production
Update in
-
Context-dependent activation of a social behavior brain network during learned vocal production.Brain Struct Funct. 2023 Sep;228(7):1785-1797. doi: 10.1007/s00429-023-02693-0. Epub 2023 Aug 24. Brain Struct Funct. 2023. PMID: 37615758
Abstract
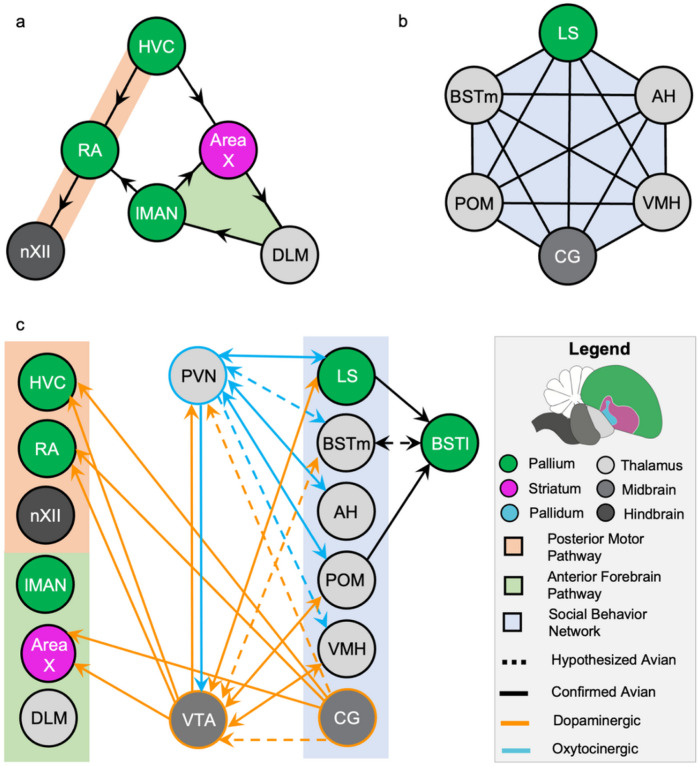
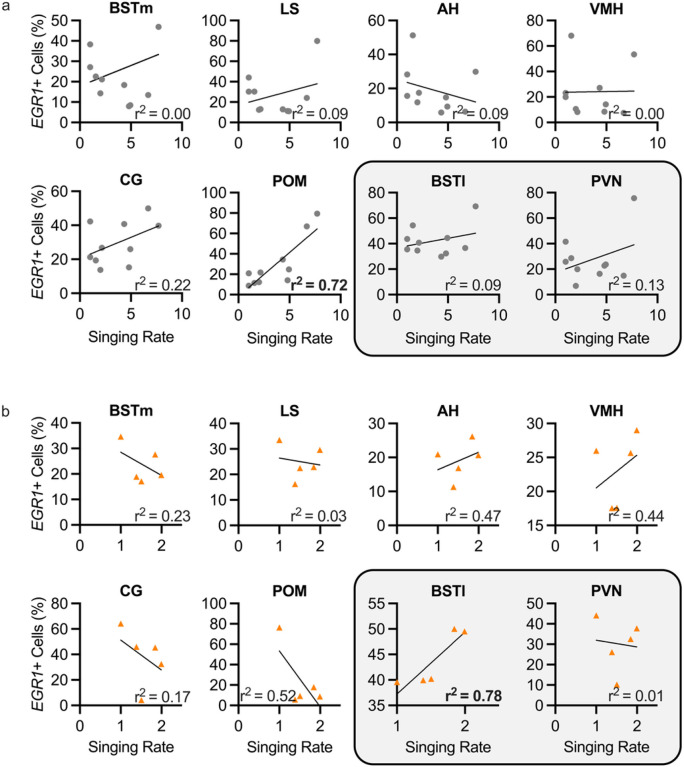
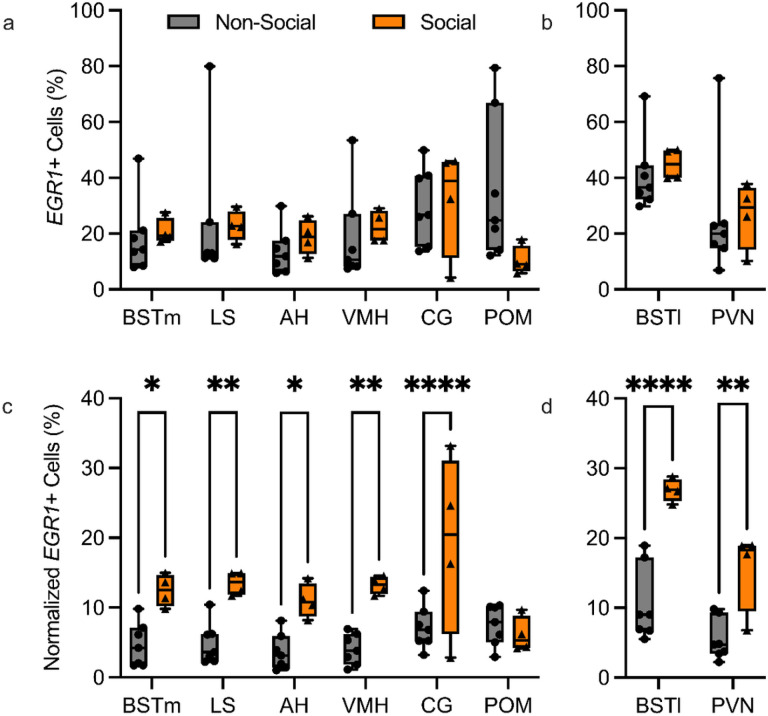
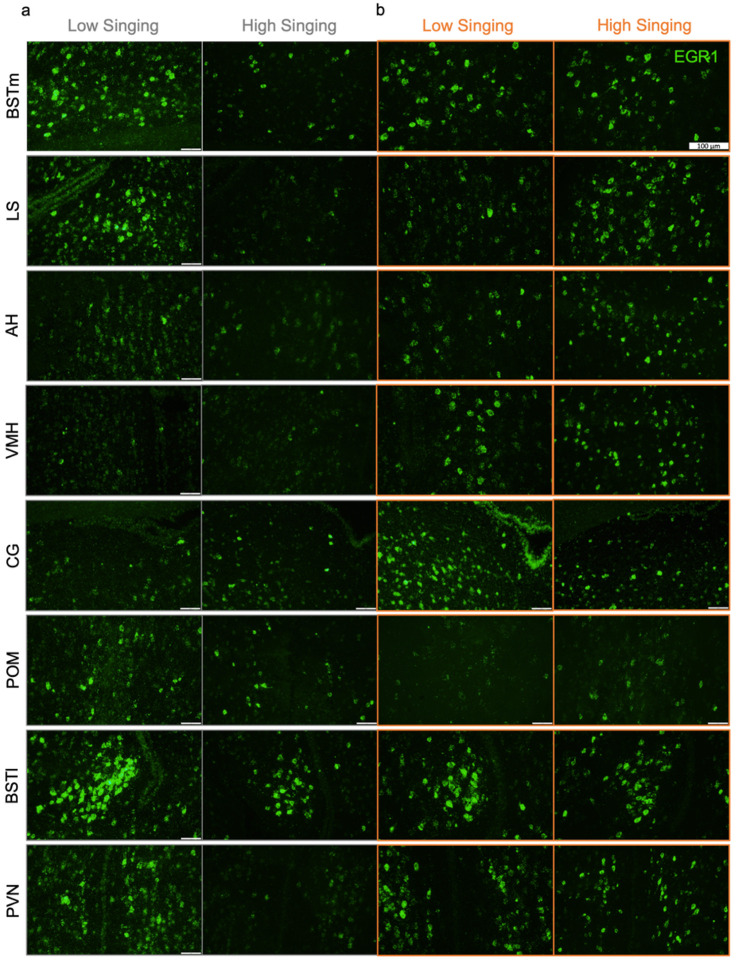
In zebra finches, an avian brain network for vocal control undergoes context-dependent patterning of song-dependent activation. Previous studies in zebra finches also implicate the importance of dopaminergic input in producing context-appropriate singing behavior. In mice, it has been shown that oxytocinergic neurons originated in the paraventricular nucleus of the hypothalamus (PVN) synapse directly onto dopamine neurons in the ventral tegmental area (VTA), implicating the necessity of oxytocin signaling from the PVN for producing a context-appropriate song. Both avian and non-avian axonal tract-tracing studies indicate high levels of PVN innervation by the social behavior network. Here, we hypothesize that the motivation for PVN oxytocin neurons to trigger dopamine release originates in the social behavior network, a highly conserved and interconnected collection of six regions implicated in various social and homeostatic behaviors. We found that expression of the neuronal activity marker EGR1 was not strongly correlated with song production in any of the regions of the social behavior network. However, when EGR1 expression levels were normalized to the singing rate, we found significantly higher levels of expression in the social behavior network regions except the medial preoptic area during a social female-directed singing context compared to a non-social undirected singing context. Our results suggest neuronal activity within the male zebra finch social behavior network influences the synaptic release of oxytocin from PVN onto dopaminergic projection neurons in the VTA, which in turn signals to the vocal control network to allow for context-appropriate song production.
Conflict of interest statement
Competing Interests
The authors have no relevant financial or non-financial conflicts of interest to disclose.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
