

This is a preprint.
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans
- PMID: 36824965
- PMCID: PMC9949080
- DOI: 10.1101/2023.02.14.528376
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans
Update in
-
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans.Cell Genom. 2024 Apr 10;4(4):100536. doi: 10.1016/j.xgen.2024.100536. Cell Genom. 2024. PMID: 38604126 Free PMC article.
Abstract
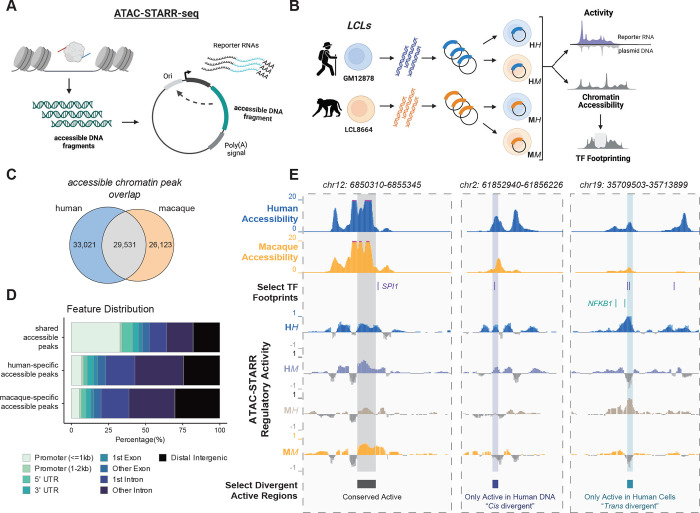
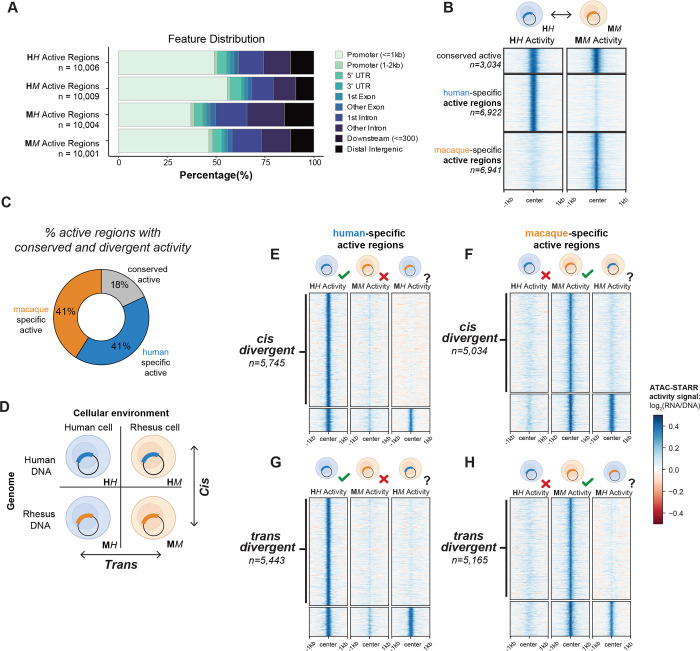
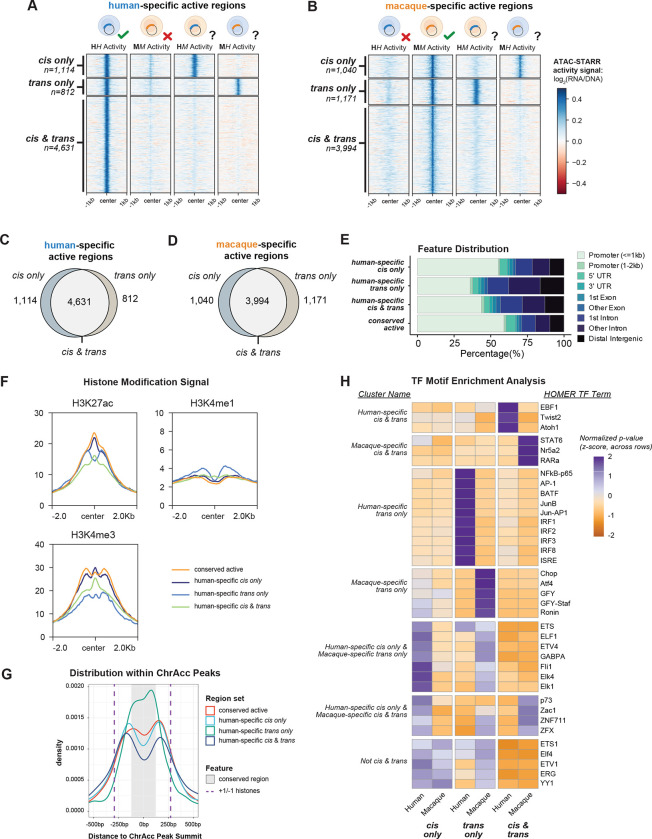
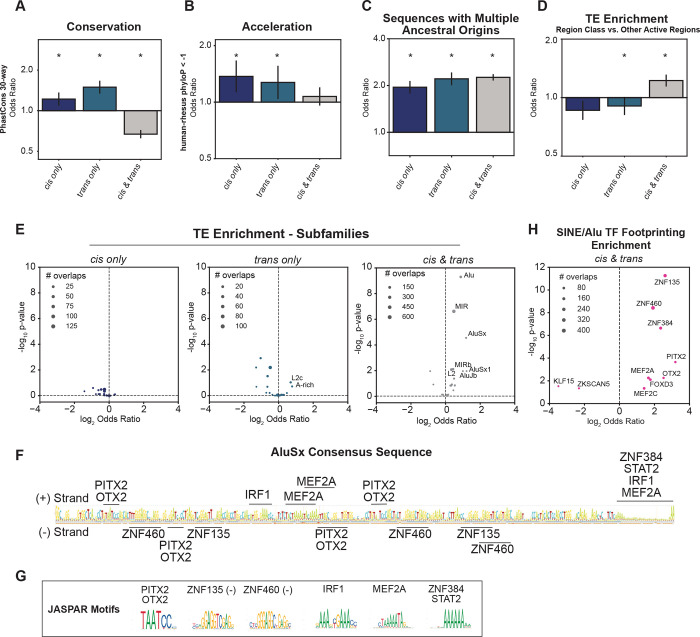
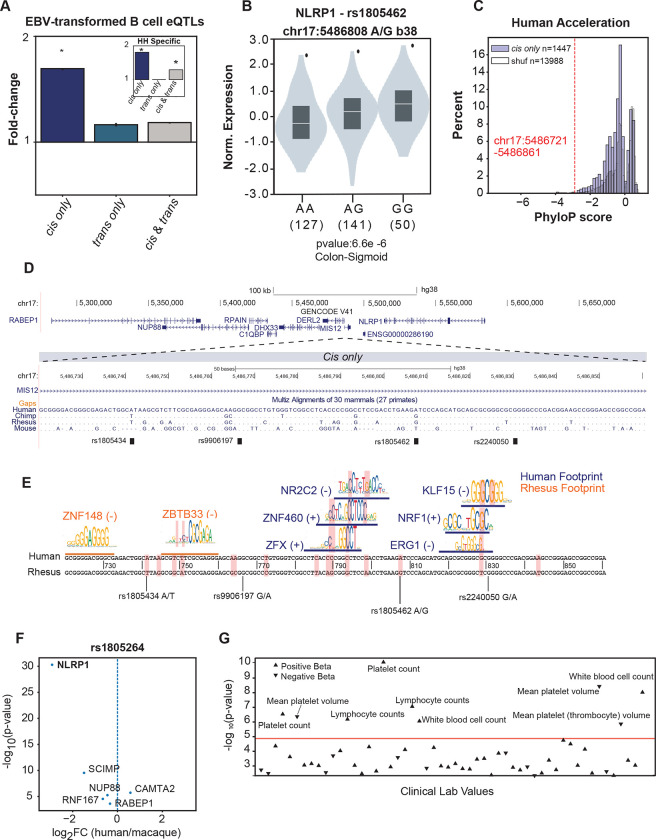
Gene regulatory divergence between species can result from cis-acting local changes to regulatory element DNA sequences or global trans-acting changes to the regulatory environment. Understanding how these mechanisms drive regulatory evolution has been limited by challenges in identifying trans-acting changes. We present a comprehensive approach to directly identify cis- and trans-divergent regulatory elements between human and rhesus macaque lymphoblastoid cells using ATAC-STARR-seq. In addition to thousands of cis changes, we discover an unexpected number (~10,000) of trans changes and show that cis and trans elements exhibit distinct patterns of sequence divergence and function. We further identify differentially expressed transcription factors that underlie >50% of trans differences and trace how cis changes can produce cascades of trans changes. Overall, we find that most divergent elements (67%) experienced changes in both cis and trans, revealing a substantial role for trans divergence-alone and together with cis changes-to regulatory differences between species.
Keywords: Comparative Genomics; DNA Regulatory Elements; Functional Genomics; Gene Regulation; Human Evolution; Lymphoblastoid Cell Lines; Massively Parallel Reporter Assays.
Conflict of interest statement
DECLERATIONS OF INTEREST The authors declare no competing interests.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials