

Ubiquitin and its relatives as wizards of the endolysosomal system
- PMID: 36825571
- PMCID: PMC10022685
- DOI: 10.1242/jcs.260101
Ubiquitin and its relatives as wizards of the endolysosomal system
Abstract
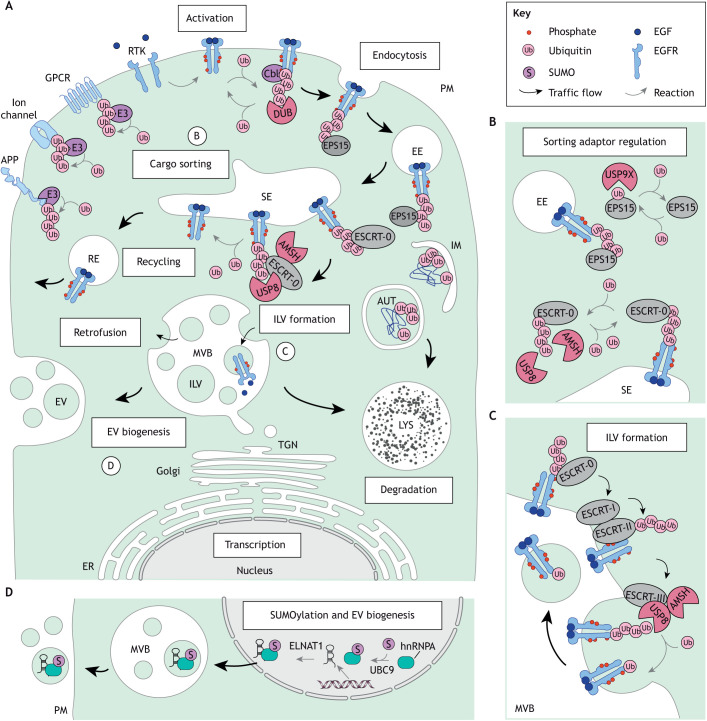
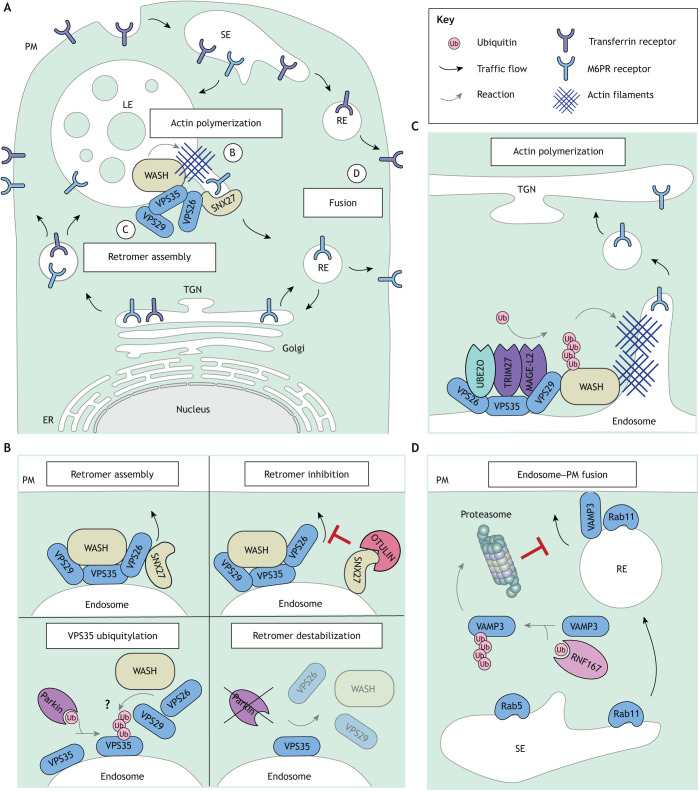
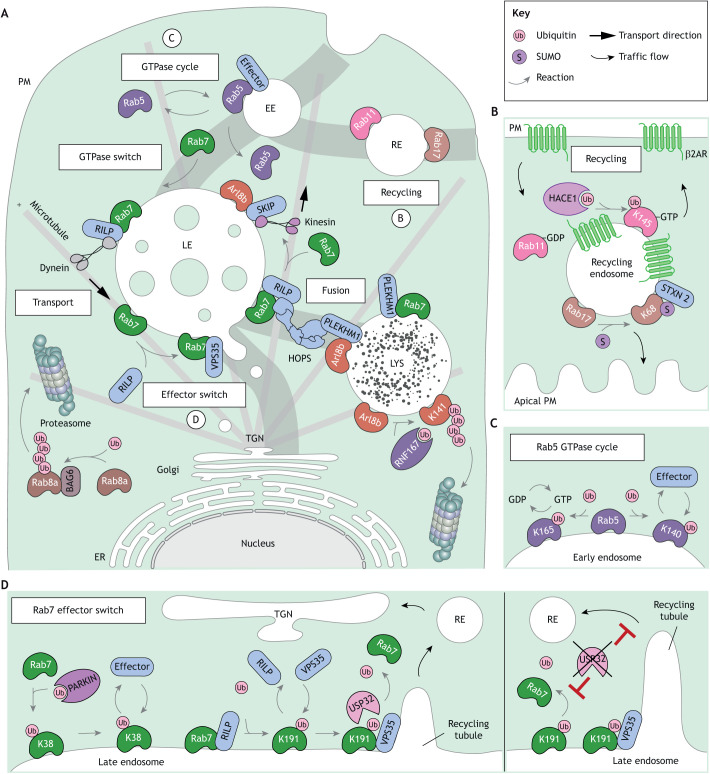
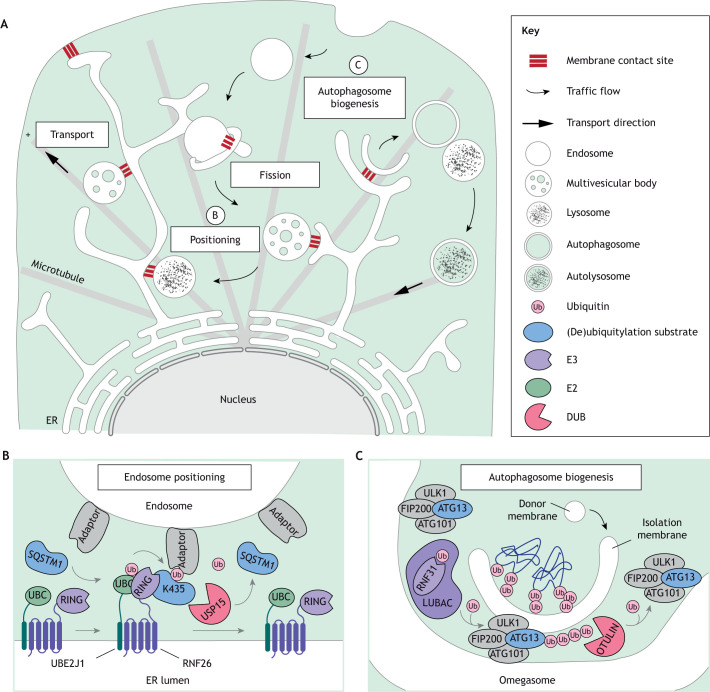
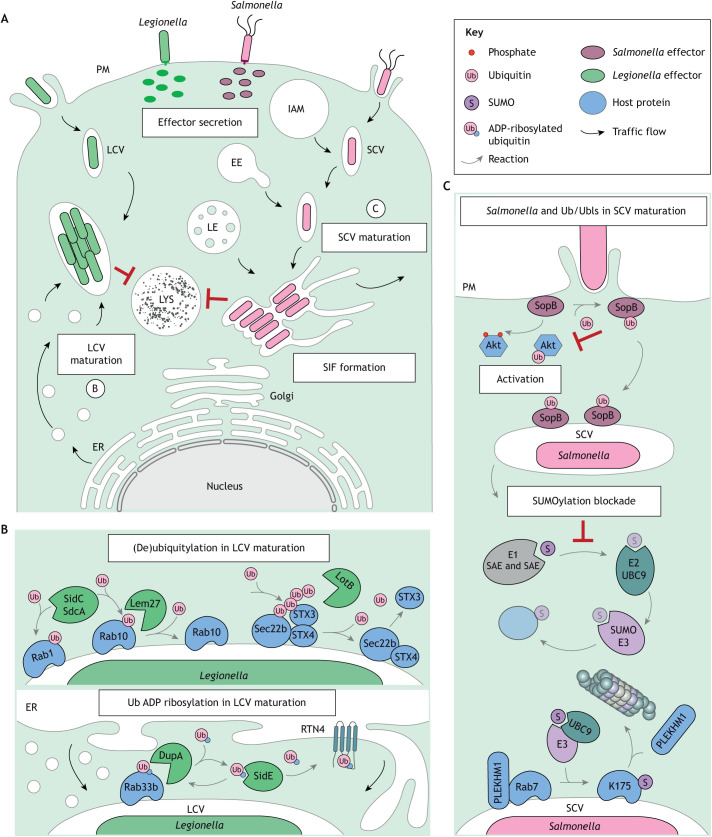
The endolysosomal system comprises a dynamic constellation of vesicles working together to sense and interpret environmental cues and facilitate homeostasis. Integrating extracellular information with the internal affairs of the cell requires endosomes and lysosomes to be proficient in decision-making: fusion or fission; recycling or degradation; fast transport or contacts with other organelles. To effectively discriminate between these options, the endolysosomal system employs complex regulatory strategies that crucially rely on reversible post-translational modifications (PTMs) with ubiquitin (Ub) and ubiquitin-like (Ubl) proteins. The cycle of conjugation, recognition and removal of different Ub- and Ubl-modified states informs cellular protein stability and behavior at spatial and temporal resolution and is thus well suited to finetune macromolecular complex assembly and function on endolysosomal membranes. Here, we discuss how ubiquitylation (also known as ubiquitination) and its biochemical relatives orchestrate endocytic traffic and designate cargo fate, influence membrane identity transitions and support formation of membrane contact sites (MCSs). Finally, we explore the opportunistic hijacking of Ub and Ubl modification cascades by intracellular bacteria that remodel host trafficking pathways to invade and prosper inside cells.
Keywords: Bacterial infection; Endosomes; Membrane contact sites; Membrane dynamics; Ubiquitin.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
Strategies to Trap Enzyme-Substrate Complexes that Mimic Michaelis Intermediates During E3-Mediated Ubiquitin-Like Protein Ligation.Methods Mol Biol. 2018;1844:169-196. doi: 10.1007/978-1-4939-8706-1_12. Methods Mol Biol. 2018. PMID: 30242710 Free PMC article.
-
The integral function of the endocytic recycling compartment is regulated by RFFL-mediated ubiquitylation of Rab11 effectors.J Cell Sci. 2019 Feb 7;132(3):jcs228007. doi: 10.1242/jcs.228007. J Cell Sci. 2019. PMID: 30659120
-
The ubiquitin code and its decoding machinery in the endocytic pathway.J Biochem. 2013 Jun;153(6):497-504. doi: 10.1093/jb/mvt028. Epub 2013 Apr 5. J Biochem. 2013. PMID: 23564907 Review.
-
Mass Spectrometry-Based Characterization of Ub- and UbL-Modified Proteins.Methods Mol Biol. 2020;2051:265-276. doi: 10.1007/978-1-4939-9744-2_11. Methods Mol Biol. 2020. PMID: 31552633
-
Genetic Code Expansion Approaches to Decipher the Ubiquitin Code.Chem Rev. 2024 Oct 23;124(20):11544-11584. doi: 10.1021/acs.chemrev.4c00375. Epub 2024 Sep 23. Chem Rev. 2024. PMID: 39311880 Free PMC article. Review.
Cited by
-
Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages.Cells. 2023 Dec 14;12(24):2833. doi: 10.3390/cells12242833. Cells. 2023. PMID: 38132160 Free PMC article.
-
PSMD9 promotes the malignant progression of hepatocellular carcinoma by interacting with c-Cbl to activate EGFR signaling and recycling.J Exp Clin Cancer Res. 2024 May 14;43(1):142. doi: 10.1186/s13046-024-03062-3. J Exp Clin Cancer Res. 2024. PMID: 38745188 Free PMC article.
-
Membrane Contact Sites in Proteostasis and ER Stress Response.Contact (Thousand Oaks). 2025 Jul 28;8:25152564251363050. doi: 10.1177/25152564251363050. eCollection 2025 Jan-Dec. Contact (Thousand Oaks). 2025. PMID: 40735229 Free PMC article. Review.
-
p97/VCP Promotes the Recycling of Endocytic Cargo.Mol Biol Cell. 2023 Dec 1;34(13):ar126. doi: 10.1091/mbc.E23-06-0237. Epub 2023 Sep 27. Mol Biol Cell. 2023. PMID: 37756124 Free PMC article.
-
E3 Ubiquitin Ligase RNF13 Suppresses TLR Lysosomal Degradation by Promoting LAMP-1 Proteasomal Degradation.Adv Sci (Weinh). 2024 Aug;11(32):e2309560. doi: 10.1002/advs.202309560. Epub 2024 Jun 21. Adv Sci (Weinh). 2024. PMID: 39031743 Free PMC article.
References
-
- Akutsu, M., Dikic, I. and Bremm, A. (2016). Ubiquitin chain diversity at a glance. J. Cell Sci. 129, 875-880. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
