

Age-associated B cells are heterogeneous and dynamic drivers of autoimmunity in mice
- PMID: 36828389
- PMCID: PMC9997508
- DOI: 10.1084/jem.20221346
Age-associated B cells are heterogeneous and dynamic drivers of autoimmunity in mice
Abstract
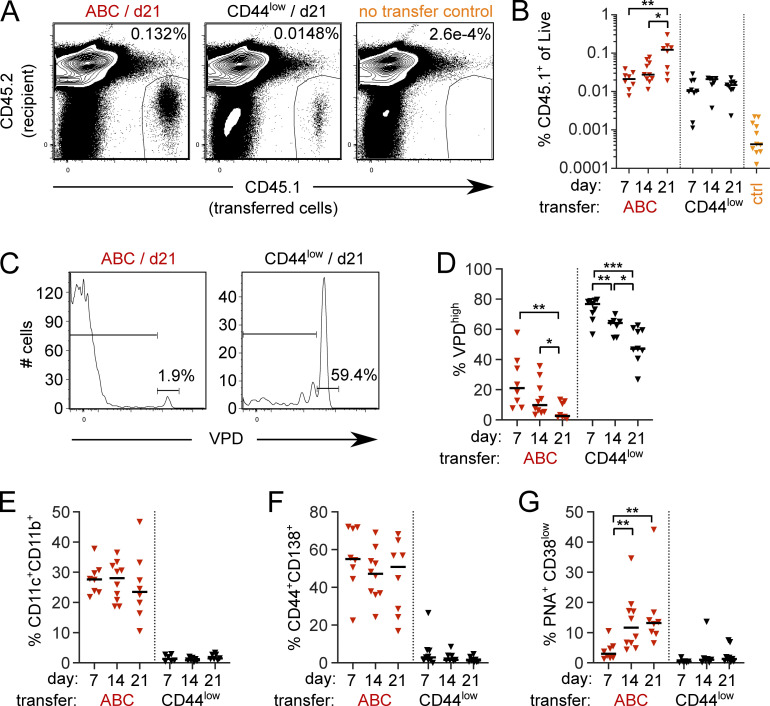
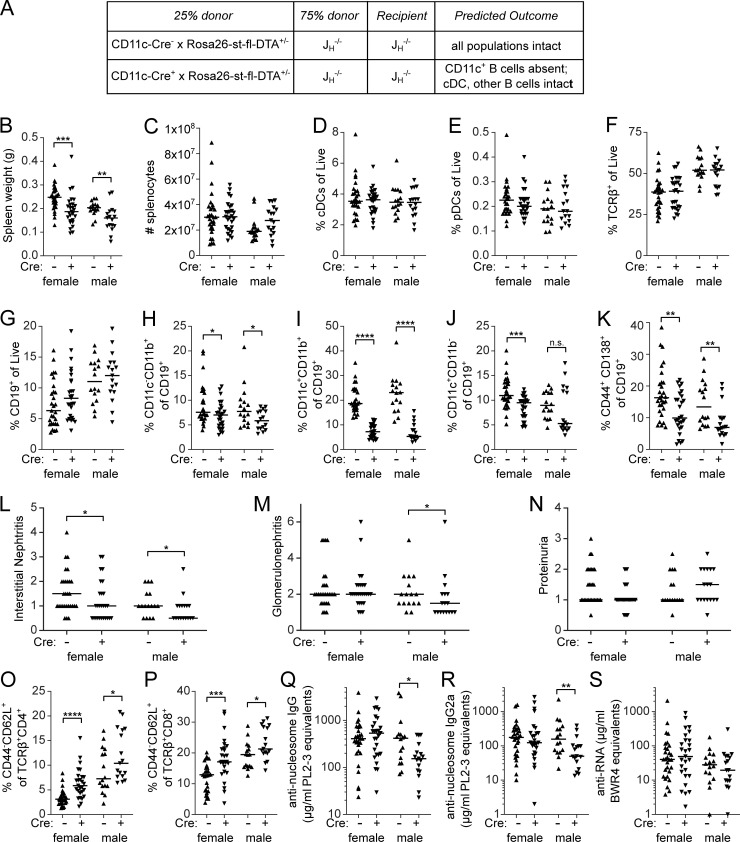
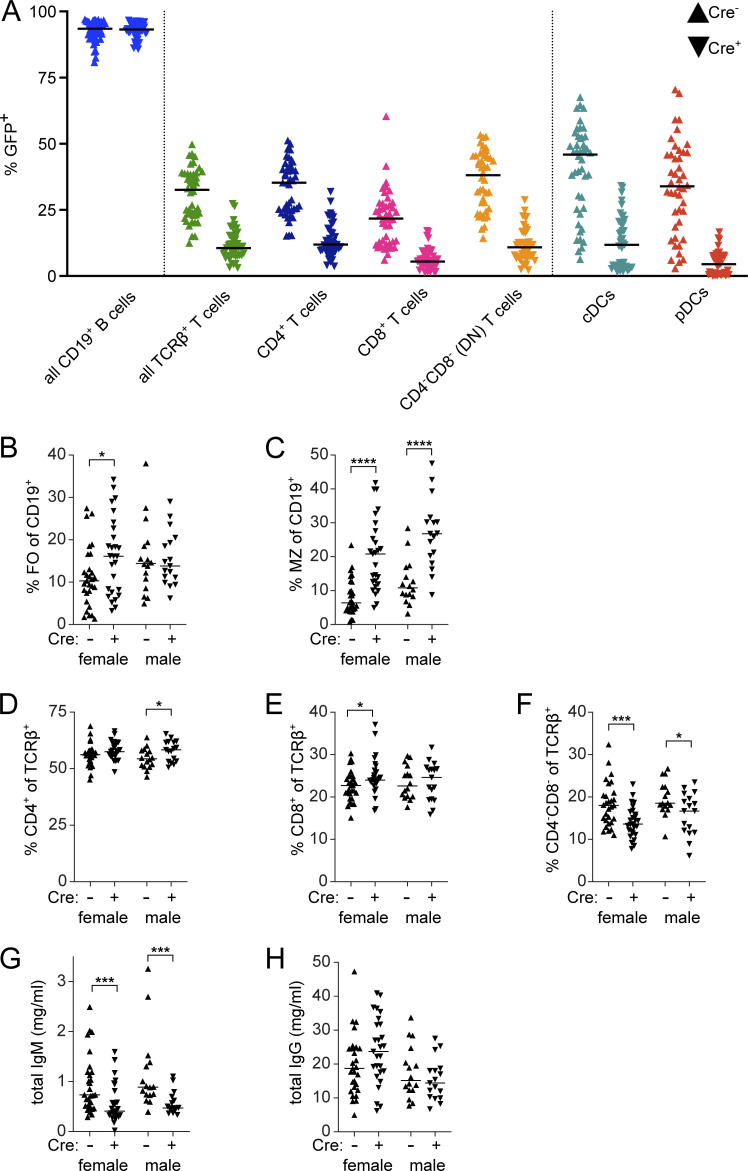
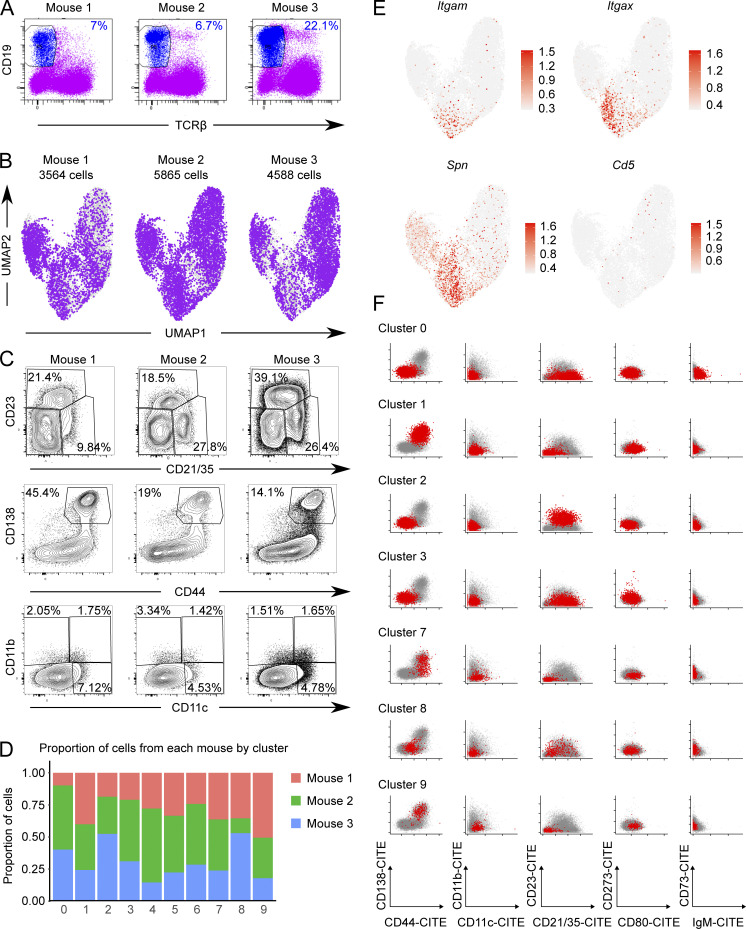
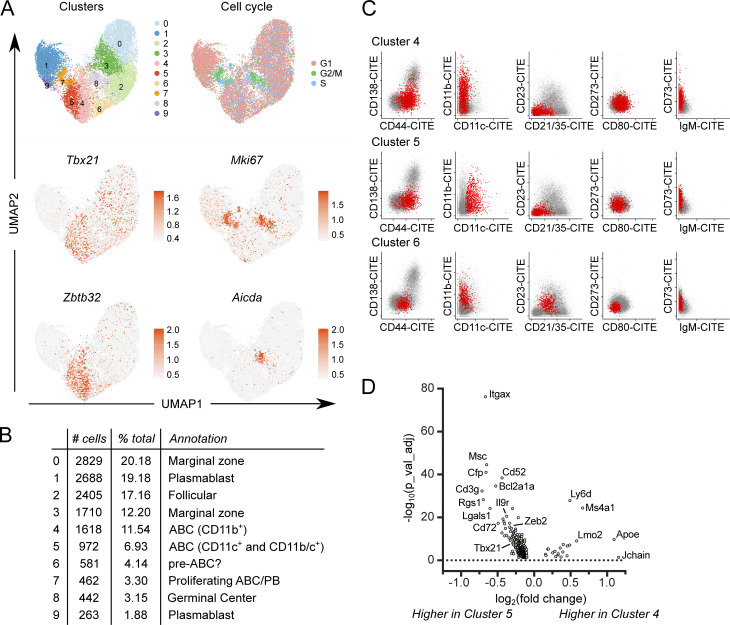
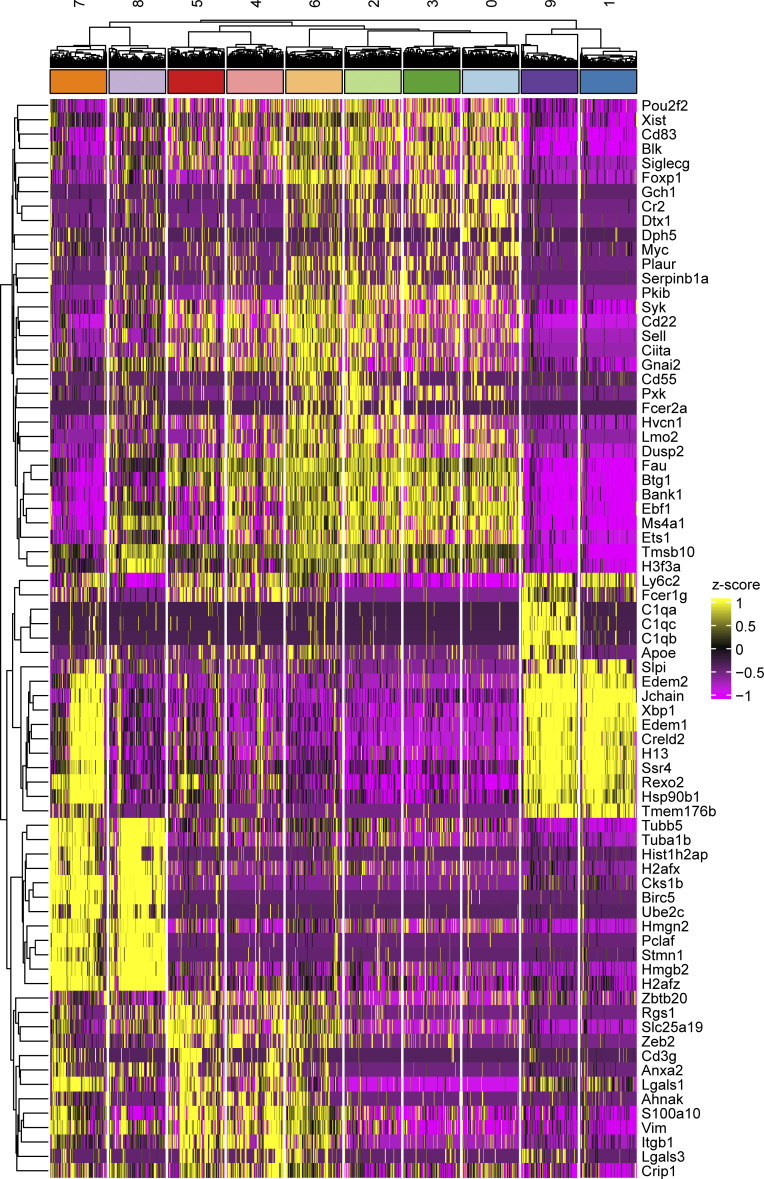
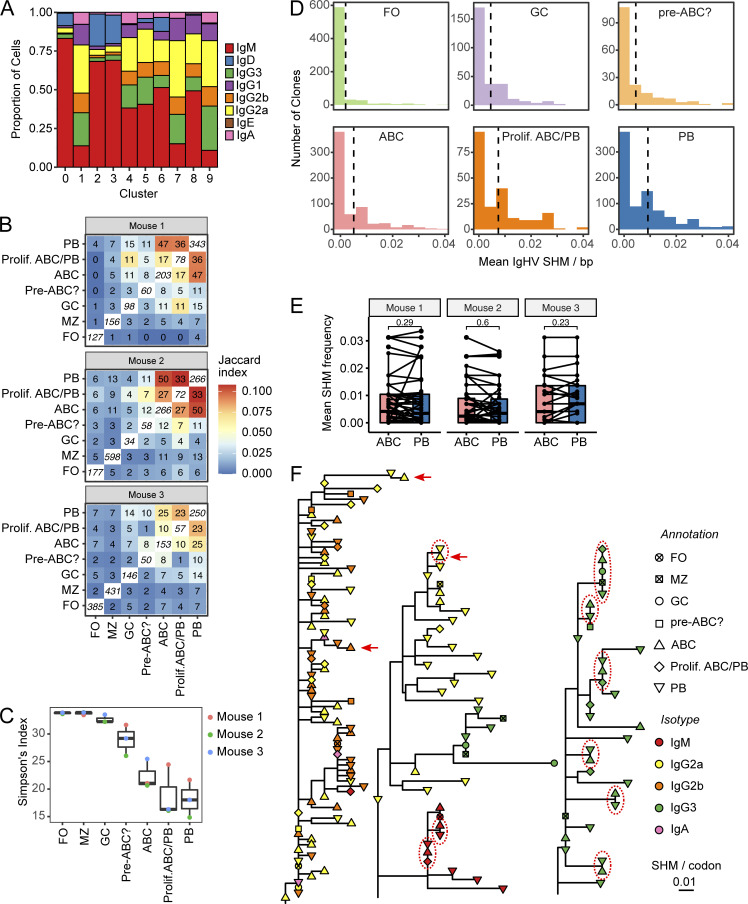
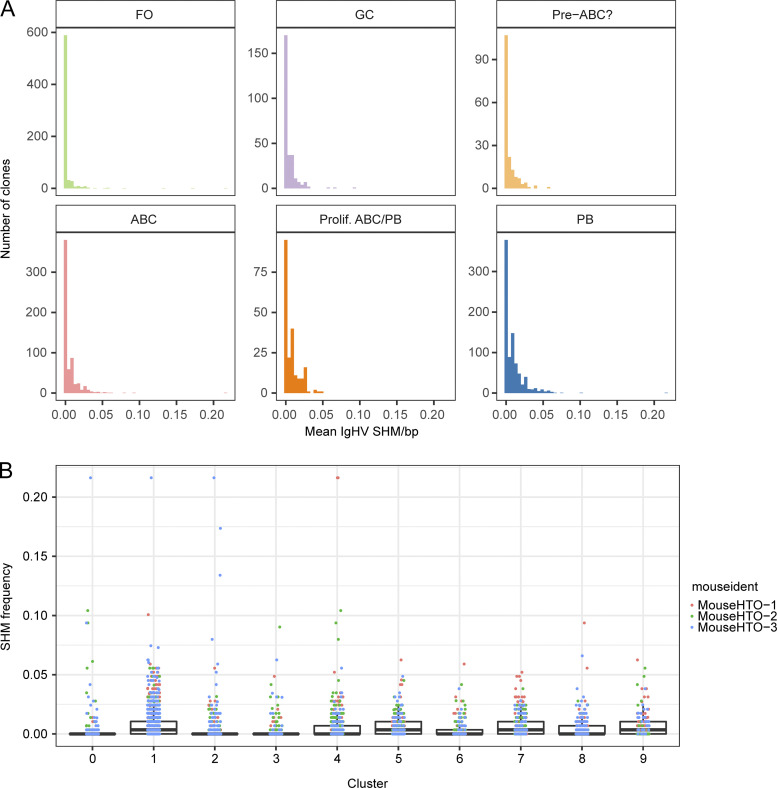
Age-associated B cells (ABCs) are formed under inflammatory conditions and are considered a type of memory B cell (MBC) expressing the transcription factor T-bet. In SLE, ABC frequency is correlated with disease, and they are thought to be the source of autoantibody-secreting cells. However, in inflammatory conditions, whether autoreactive B cells can become resting MBCs is uncertain. Further, the phenotypic identity of ABCs and their relationship to other B cell subsets, such as plasmablasts, is unclear. Whether ABCs directly promote disease is untested. Here we report, in the MRL/lpr SLE model, unexpected heterogeneity among ABC-like cells for expression of the integrins CD11b and CD11c, T-bet, and memory or plasmablast markers. Transfer and labeling studies demonstrated that ABCs are dynamic, rapidly turning over. scRNA-seq identified B cell clones present in multiple subsets, revealing that ABCs can be plasmablast precursors or undergo cycles of reactivation. Deletion of CD11c-expressing B cells revealed a direct role for ABC-like B cells in lupus pathogenesis.
© 2023 Nickerson et al.
Conflict of interest statement
Disclosures: K.B. Hoehn reported personal fees from Prellis Biologics outside the submitted work. S.H. Kleinstein reported consulting fees from Peraton. No other disclosures were reported.
Figures
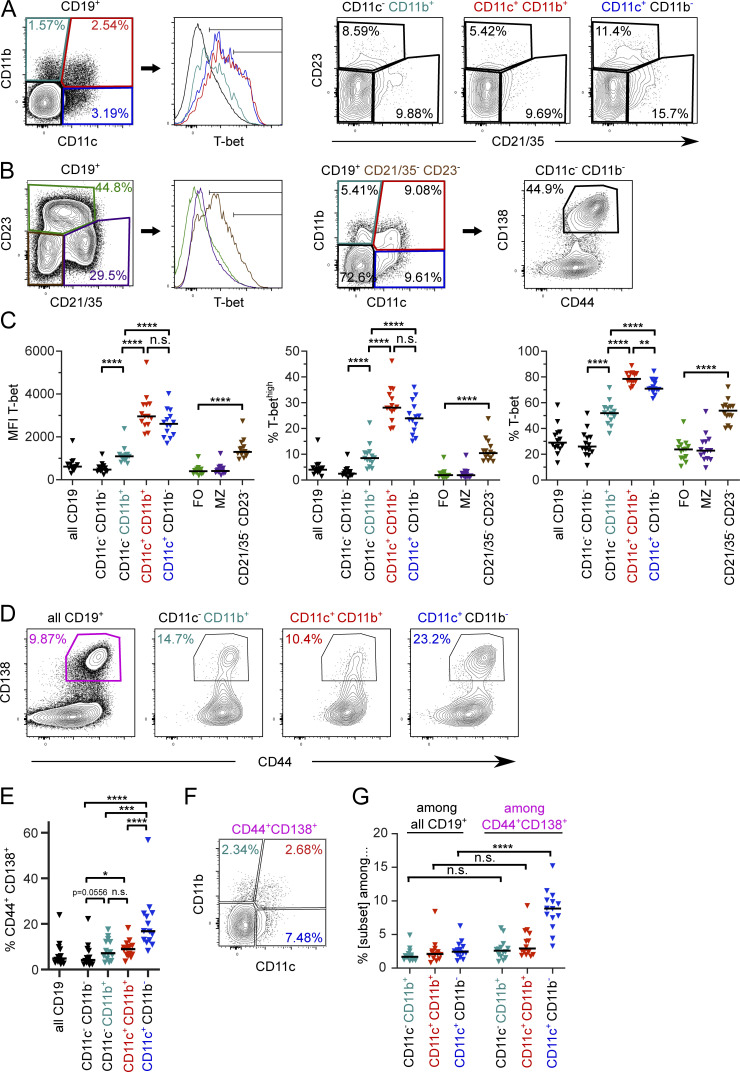
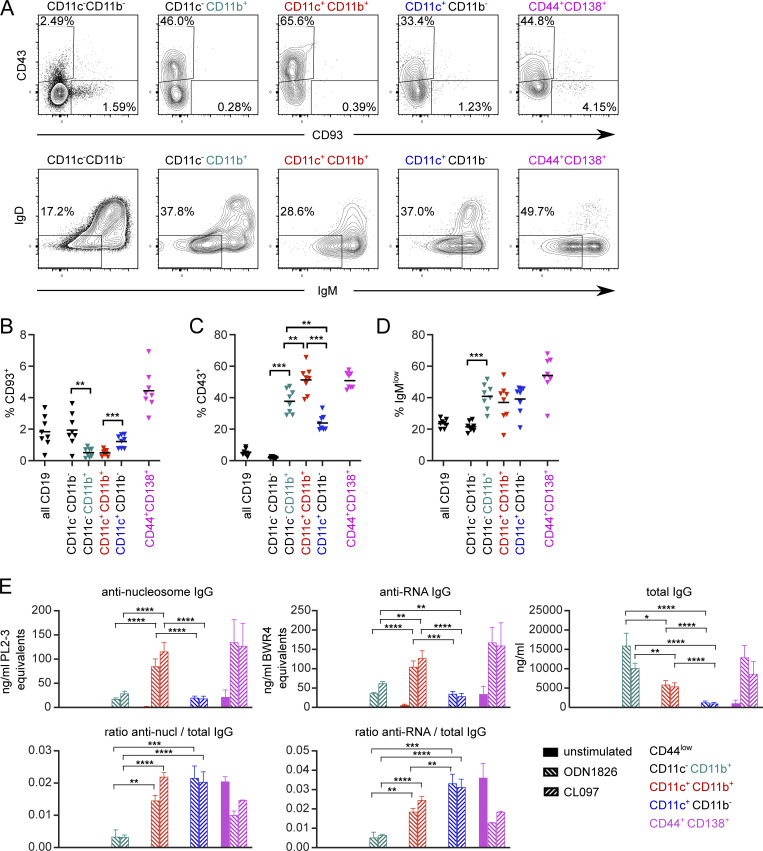

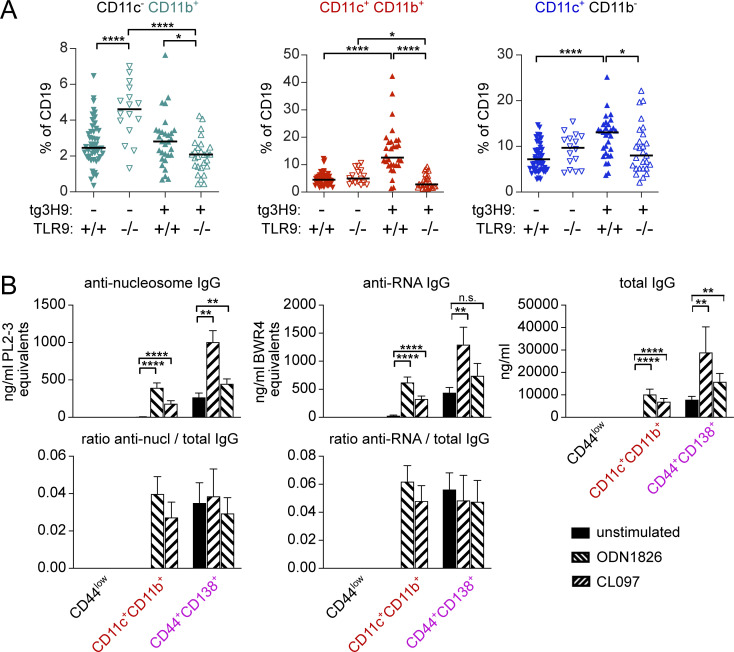
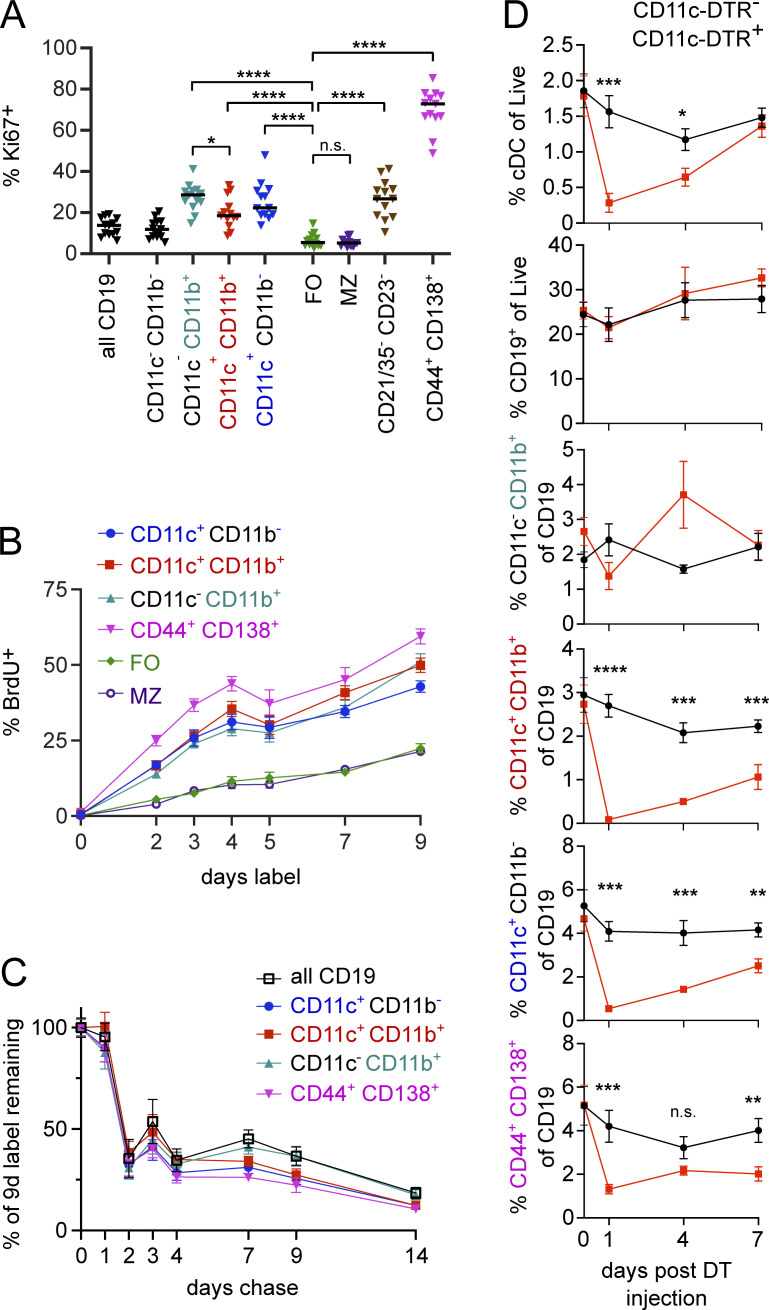
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
