

Selective Destabilization of Transcripts by mRNA Decapping Regulates Oocyte Maturation and Innate Immunity Gene Expression during Ageing in C. elegans
- PMID: 36829450
- PMCID: PMC9952881
- DOI: 10.3390/biology12020171
Selective Destabilization of Transcripts by mRNA Decapping Regulates Oocyte Maturation and Innate Immunity Gene Expression during Ageing in C. elegans
Abstract
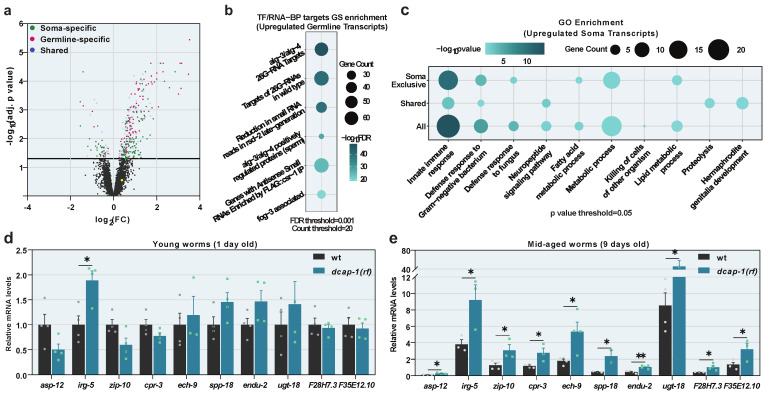
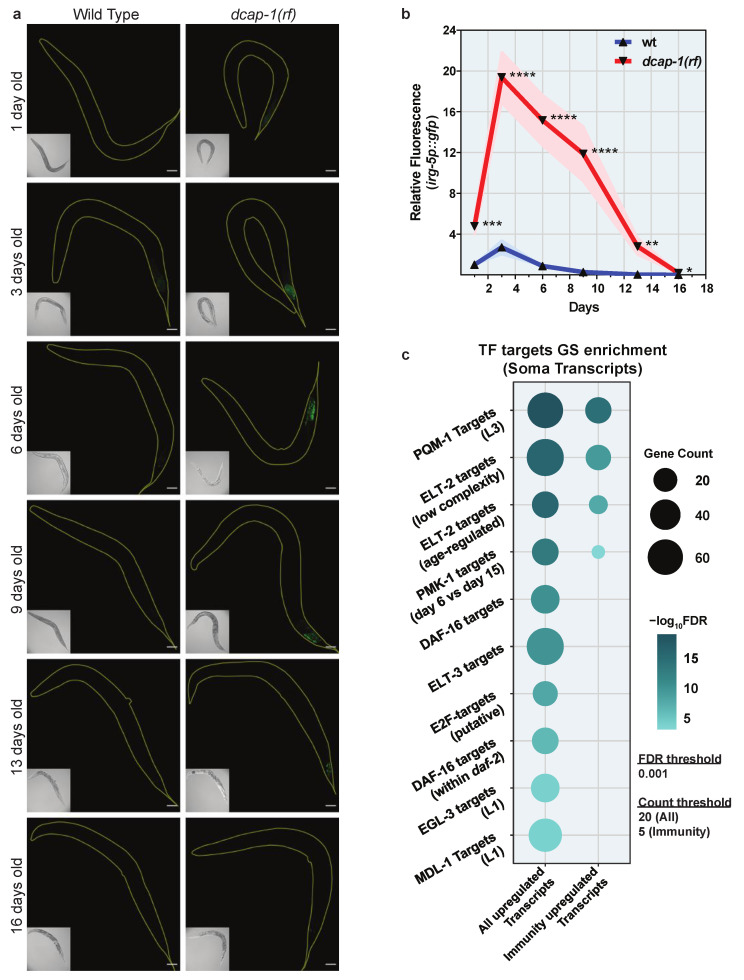
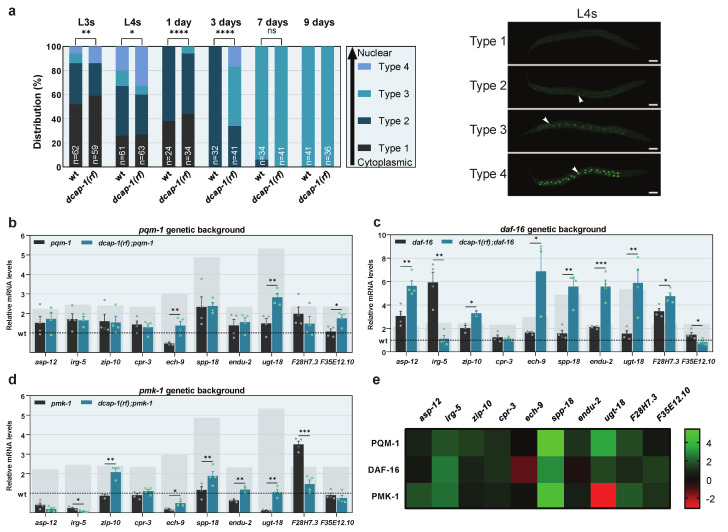
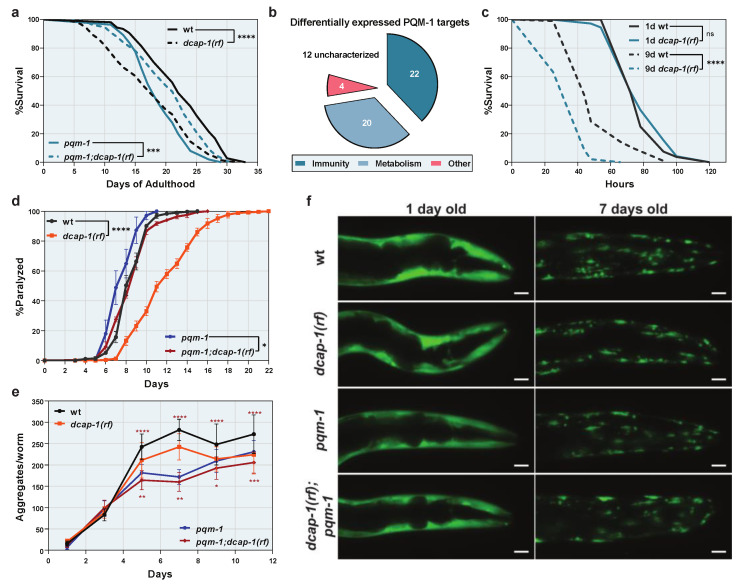
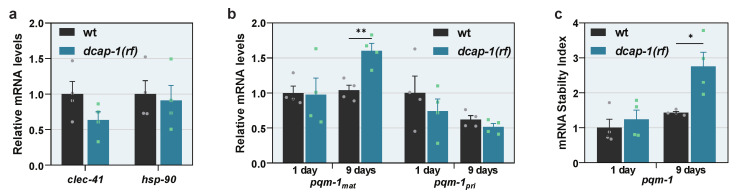
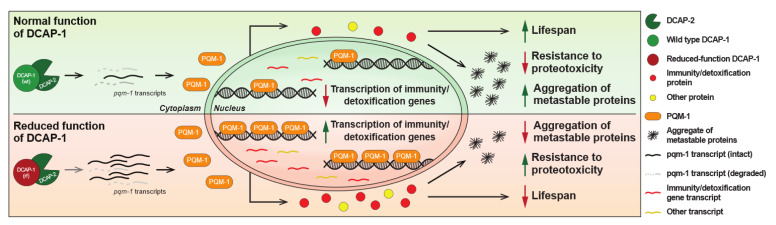
Removal of the 5' cap structure of RNAs (termed decapping) is a pivotal event in the life of cytoplasmic mRNAs mainly catalyzed by a conserved holoenzyme, composed of the catalytic subunit DCP2 and its essential cofactor DCP1. While decapping was initially considered merely a step in the general 5'-3' mRNA decay, recent data suggest a great degree of selectivity that plays an active role in the post-transcriptional control of gene expression, and regulates multiple biological functions. Studies in Caenorhabditis elegans have shown that old age is accompanied by the accumulation of decapping factors in cytoplasmic RNA granules, and loss of decapping activity shortens the lifespan. However, the link between decapping and ageing remains elusive. Here, we present a comparative microarray study that was aimed to uncover the differences in the transcriptome of mid-aged dcap-1/DCP1 mutant and wild-type nematodes. Our data indicate that DCAP-1 mediates the silencing of spermatogenic genes during late oogenesis, and suppresses the aberrant uprise of immunity gene expression during ageing. The latter is achieved by destabilizing the mRNA that encodes the transcription factor PQM-1 and impairing its nuclear translocation. Failure to exert decapping-mediated control on PQM-1 has a negative impact on the lifespan, but mitigates the toxic effects of polyglutamine expression that are involved in human disease.
Keywords: C. elegans; DCAP-1; PQM-1; ageing; innate immunity; mRNA decapping; microarray; polyglutamine; spermatogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
