

Updated Virophage Taxonomy and Distinction from Polinton-like Viruses
- PMID: 36830574
- PMCID: PMC9952930
- DOI: 10.3390/biom13020204
Updated Virophage Taxonomy and Distinction from Polinton-like Viruses
Abstract
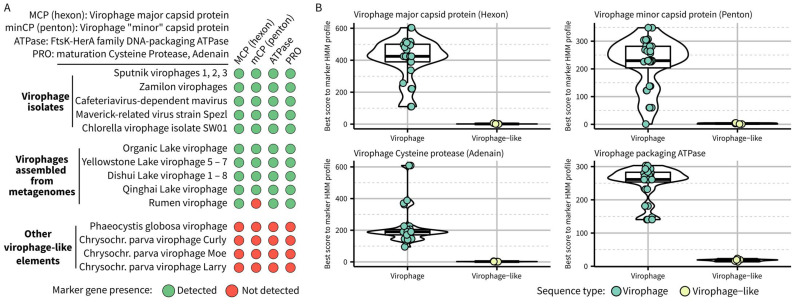
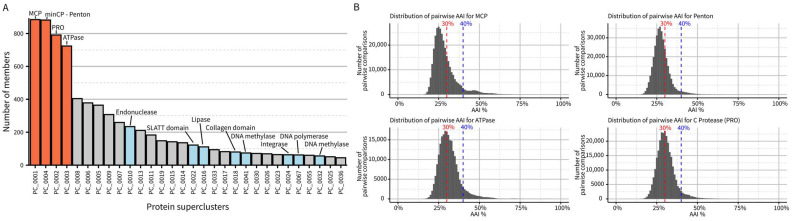
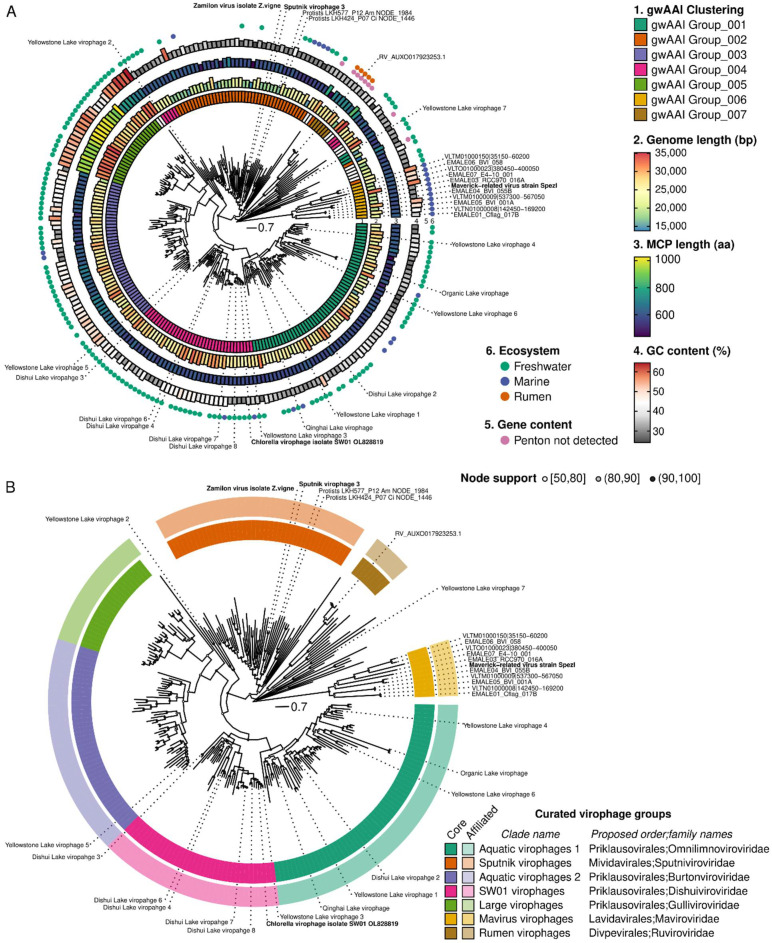
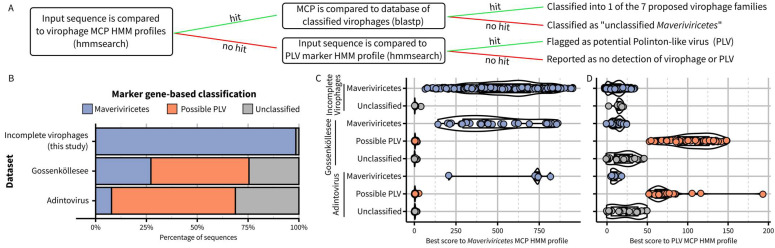
Virophages are small dsDNA viruses that hijack the machinery of giant viruses during the co-infection of a protist (i.e., microeukaryotic) host and represent an exceptional case of "hyperparasitism" in the viral world. While only a handful of virophages have been isolated, a vast diversity of virophage-like sequences have been uncovered from diverse metagenomes. Their wide ecological distribution, idiosyncratic infection and replication strategy, ability to integrate into protist and giant virus genomes and potential role in antiviral defense have made virophages a topic of broad interest. However, one limitation for further studies is the lack of clarity regarding the nomenclature and taxonomy of this group of viruses. Specifically, virophages have been linked in the literature to other "virophage-like" mobile genetic elements and viruses, including polinton-like viruses (PLVs), but there are no formal demarcation criteria and proper nomenclature for either group, i.e., virophage or PLVs. Here, as part of the ICTV Virophage Study Group, we leverage a large set of genomes gathered from published datasets as well as newly generated protist genomes to propose delineation criteria and classification methods at multiple taxonomic ranks for virophages 'sensu stricto', i.e., genomes related to the prototype isolates Sputnik and mavirus. Based on a combination of comparative genomics and phylogenetic analyses, we show that this group of virophages forms a cohesive taxon that we propose to establish at the class level and suggest a subdivision into four orders and seven families with distinctive ecogenomic features. Finally, to illustrate how the proposed delineation criteria and classification method would be used, we apply these to two recently published datasets, which we show include both virophages and other virophage-related elements. Overall, we see this proposed classification as a necessary first step to provide a robust taxonomic framework in this area of the virosphere, which will need to be expanded in the future to cover other virophage-related viruses such as PLVs.
Keywords: giant virus; polinton; polintovirus; virophage; virus taxonomy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Isolation and Identification of a Large Green Alga Virus (Chlorella Virus XW01) of Mimiviridae and Its Virophage (Chlorella Virus Virophage SW01) by Using Unicellular Green Algal Cultures.J Virol. 2022 Apr 13;96(7):e0211421. doi: 10.1128/jvi.02114-21. Epub 2022 Mar 9. J Virol. 2022. PMID: 35262372 Free PMC article.
-
Virophages and retrotransposons colonize the genomes of a heterotrophic flagellate.Elife. 2021 Oct 26;10:e72674. doi: 10.7554/eLife.72674. Elife. 2021. PMID: 34698016 Free PMC article.
-
Polinton-like viruses are abundant in aquatic ecosystems.Microbiome. 2021 Jan 12;9(1):13. doi: 10.1186/s40168-020-00956-0. Microbiome. 2021. PMID: 33436089 Free PMC article.
-
Polintons, virophages and transpovirons: a tangled web linking viruses, transposons and immunity.Curr Opin Virol. 2017 Aug;25:7-15. doi: 10.1016/j.coviro.2017.06.008. Epub 2017 Jun 30. Curr Opin Virol. 2017. PMID: 28672161 Free PMC article. Review.
-
The polinton-like supergroup of viruses: evolution, molecular biology, and taxonomy.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0008623. doi: 10.1128/mmbr.00086-23. Epub 2024 Jul 18. Microbiol Mol Biol Rev. 2024. PMID: 39023254 Free PMC article. Review.
Cited by
-
Lineage-specific expansions of polinton-like viruses in photosynthetic cryptophytes.Microbiome. 2025 Jul 1;13(1):154. doi: 10.1186/s40168-025-02148-0. Microbiome. 2025. PMID: 40598447 Free PMC article.
-
Genomic analysis of hyperparasitic viruses associated with entomopoxviruses.Virus Evol. 2024 Jul 12;10(1):veae051. doi: 10.1093/ve/veae051. eCollection 2024. Virus Evol. 2024. PMID: 39100687 Free PMC article.
-
Virophages-Known and Unknown Facts.Viruses. 2023 Jun 5;15(6):1321. doi: 10.3390/v15061321. Viruses. 2023. PMID: 37376621 Free PMC article. Review.
-
Astrovirology: how viruses enhance our understanding of life in the Universe.Int J Astrobiol. 2023 Aug;22(4):247-271. doi: 10.1017/s1473550423000058. Epub 2023 Apr 5. Int J Astrobiol. 2023. PMID: 38046673 Free PMC article.
-
Virophages Found in Viromes from Lake Baikal.Biomolecules. 2023 Dec 11;13(12):1773. doi: 10.3390/biom13121773. Biomolecules. 2023. PMID: 38136644 Free PMC article.
