

CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato
- PMID: 36835025
- PMCID: PMC9964222
- DOI: 10.3390/ijms24043613
CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato
Abstract
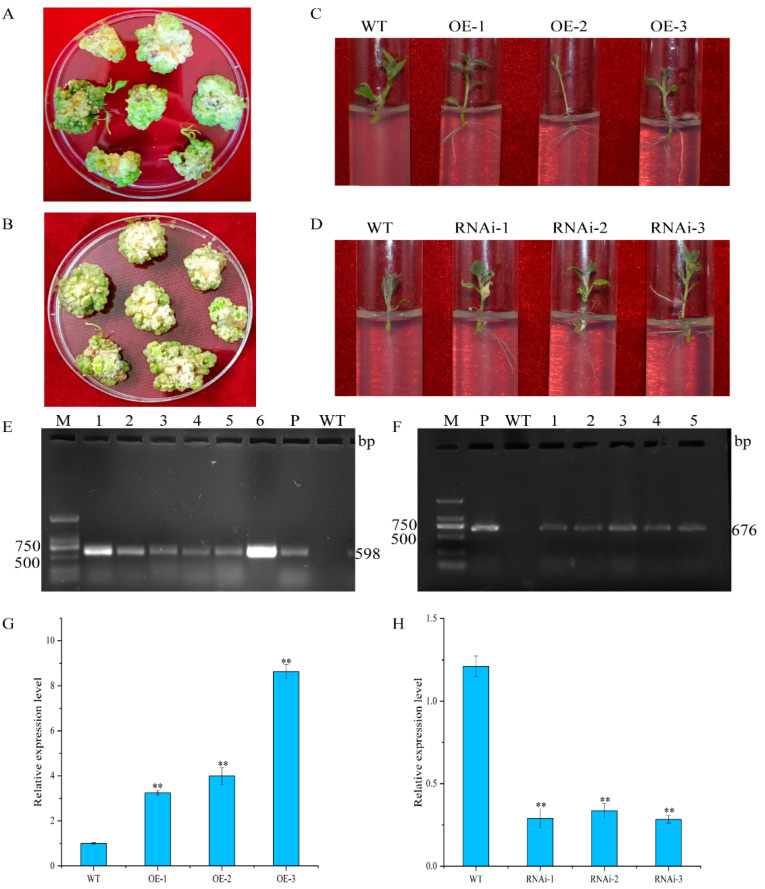
Sensor-responder complexes comprising calcineurin B-like (CBL) proteins and CBL-interacting protein kinases (CIPKs) are plant-specific Ca2+ receptors, and the CBL-CIPK module is widely involved in plant growth and development and a large number of abiotic stress response signaling pathways. In this study, the potato cv. "Atlantic" was subjected to a water deficiency treatment and the expression of StCIPK18 gene was detected by qRT-PCR. The subcellular localization of StCIPK18 protein was observed by a confocal laser scanning microscope. The StCIPK18 interacting protein was identified and verified by yeast two-hybrid (Y2H) and bimolecular fluorescence complementation (BiFC). StCIPK18 overexpression and StCIPK18 knockout plants were constructed. The phenotypic changes under drought stress were indicated by water loss rate, relative water content, MDA and proline contents, and CAT, SOD and POD activities. The results showed that StCIPK18 expression was upregulated under drought stress. StCIPK18 is localized in the cell membrane and cytoplasm. Y2H shows the interaction between StCIPK18 and StCBL1, StCBL4, StCBL6 and StCBL8. BiFC further verifies the reliability of the interaction between StCIPK18 and StCBL4. Under drought stress, StCIPK18 overexpression decreased the water loss rate and MDA, and increased RWC, proline contents and CAT, SOD and POD activities; however, StCIPK18 knockout showed opposite results, compared with the wild type, in response to drought stress. The results can provide information for the molecular mechanism of the StCIPK18 regulating potato response to drought stress.
Keywords: StCIPK18; drought stress; potato; protein interaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Sheng J., Zheng Z., Pan W., Pan J. Functions and action Mechanisms of CBL-CIPK signaling system in plants. Plant Physiol. J. 2014;50:641–650.
-
- Ma R., Li S., Liu W., Wang Z., Yang J., Tang X., Zhang N., Si H. Functions and progress in mechanism research of CBL-CIPK signaling system in plants. Plant Physiol. J. 2021;57:521–530.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
