

Responses of Aerial and Belowground Parts of Different Potato (Solanum tuberosum L.) Cultivars to Heat Stress
- PMID: 36840167
- PMCID: PMC9964869
- DOI: 10.3390/plants12040818
Responses of Aerial and Belowground Parts of Different Potato (Solanum tuberosum L.) Cultivars to Heat Stress
Abstract
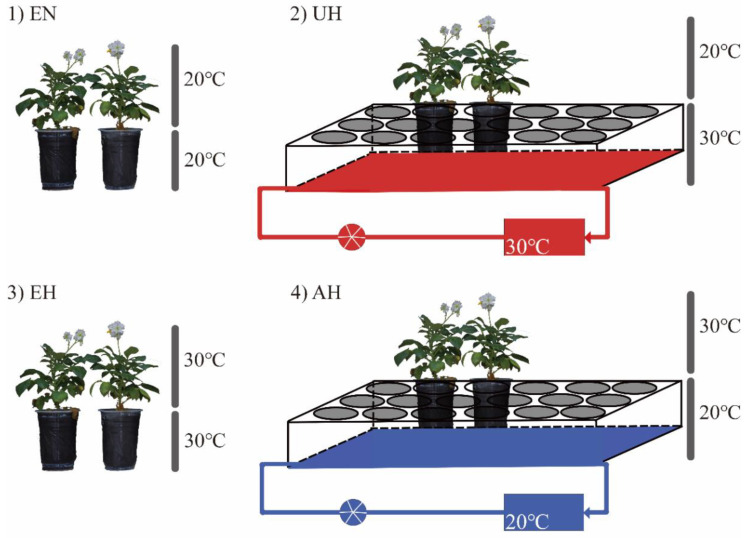
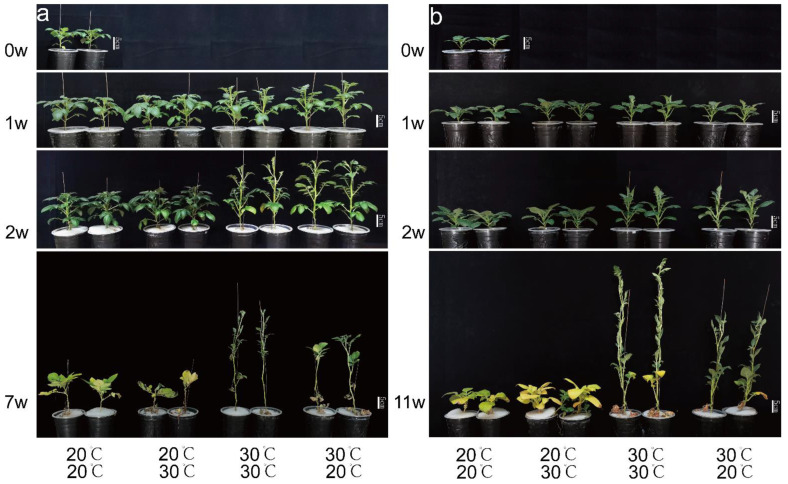
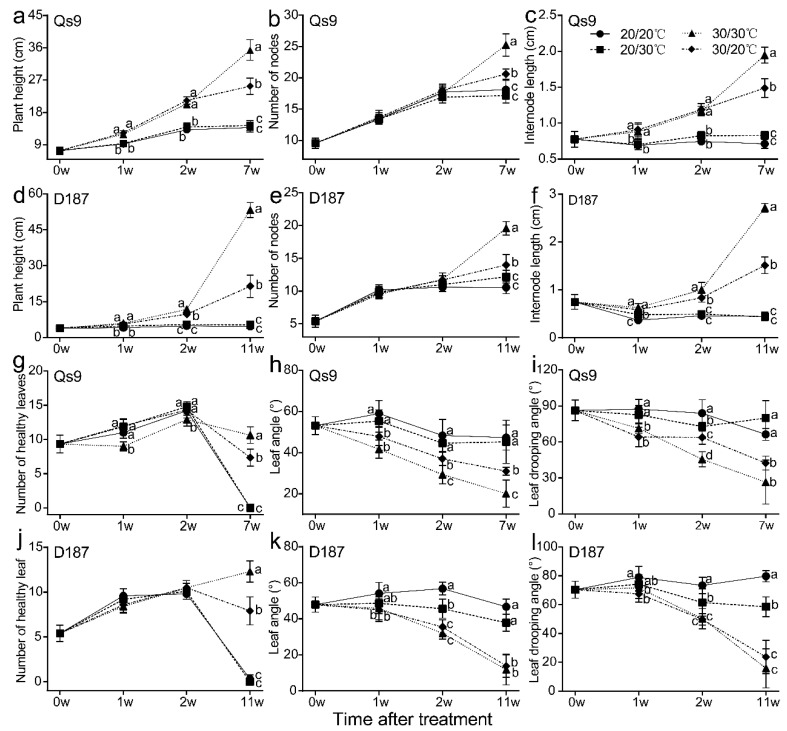
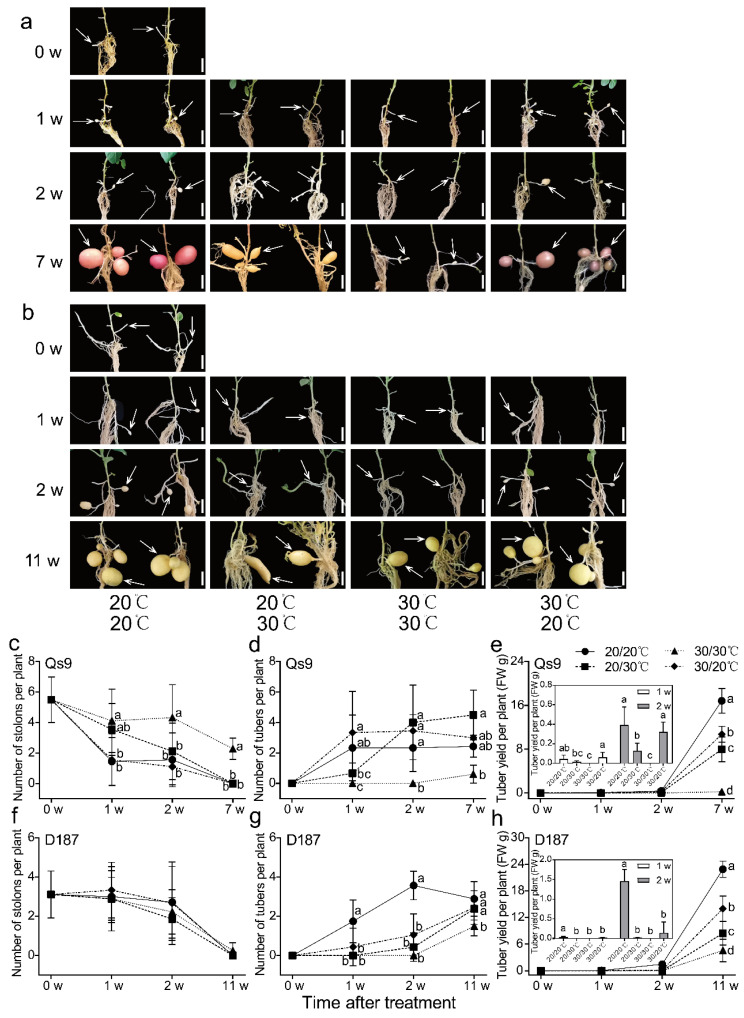
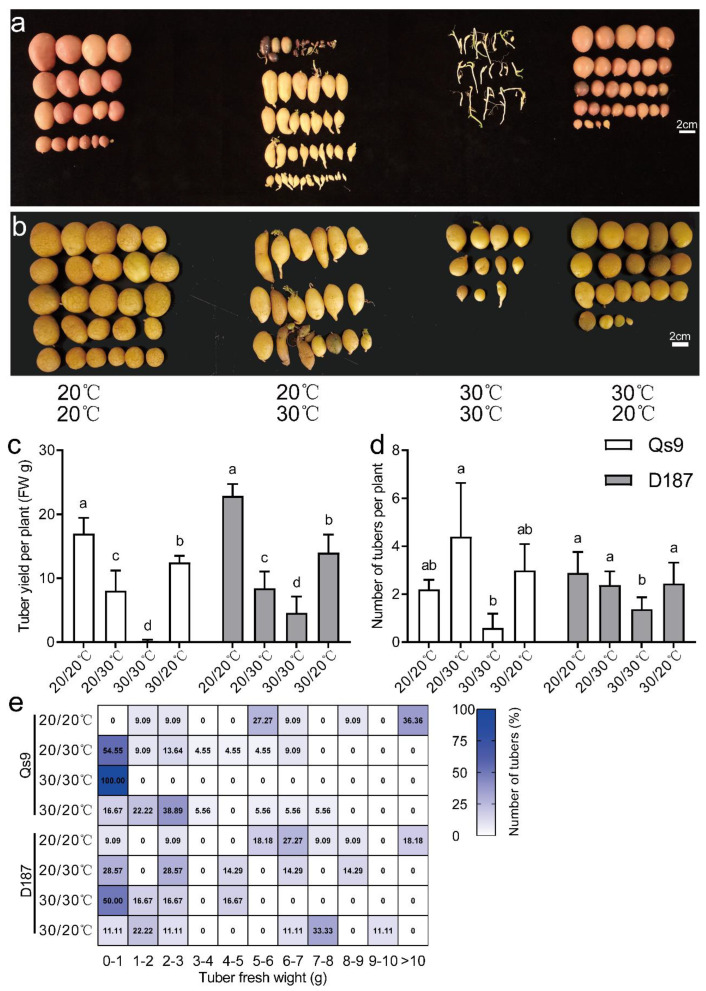
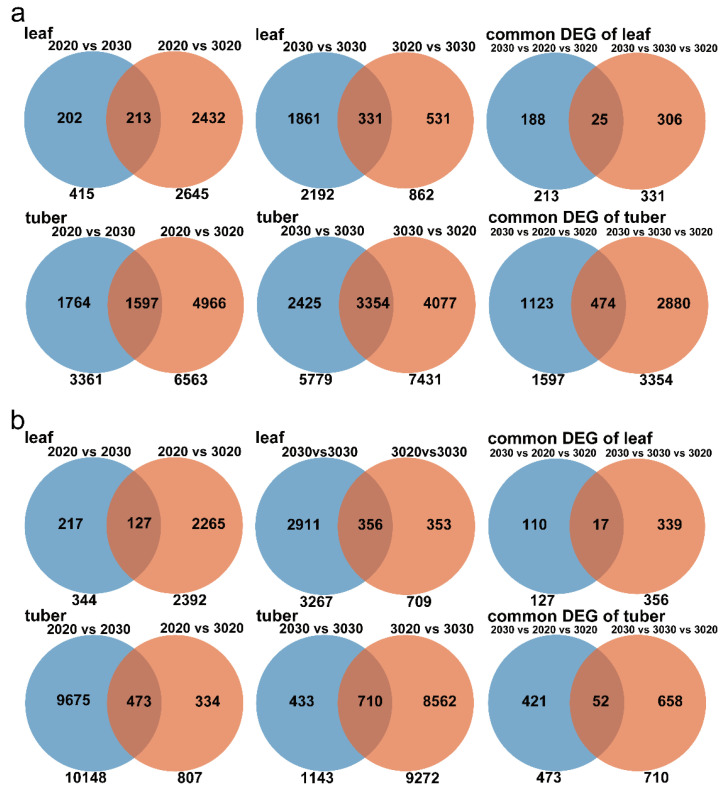
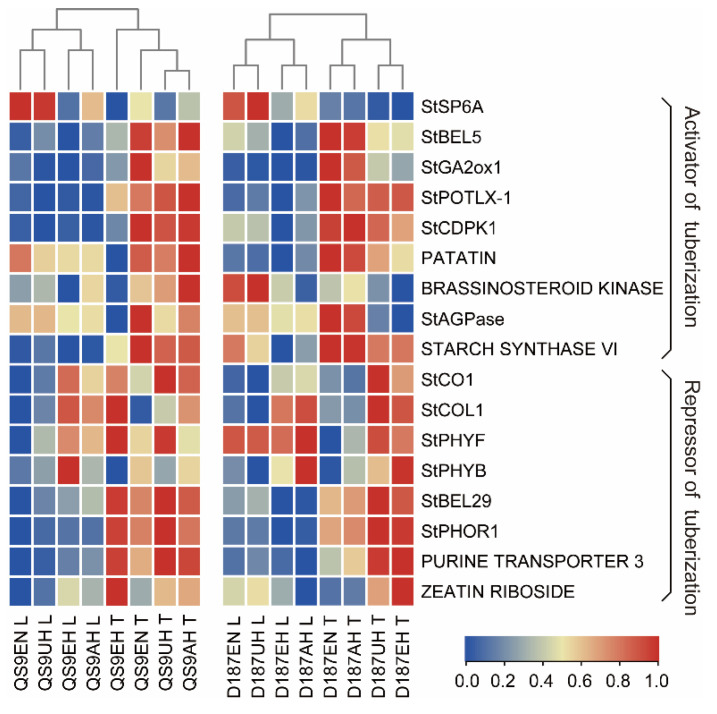
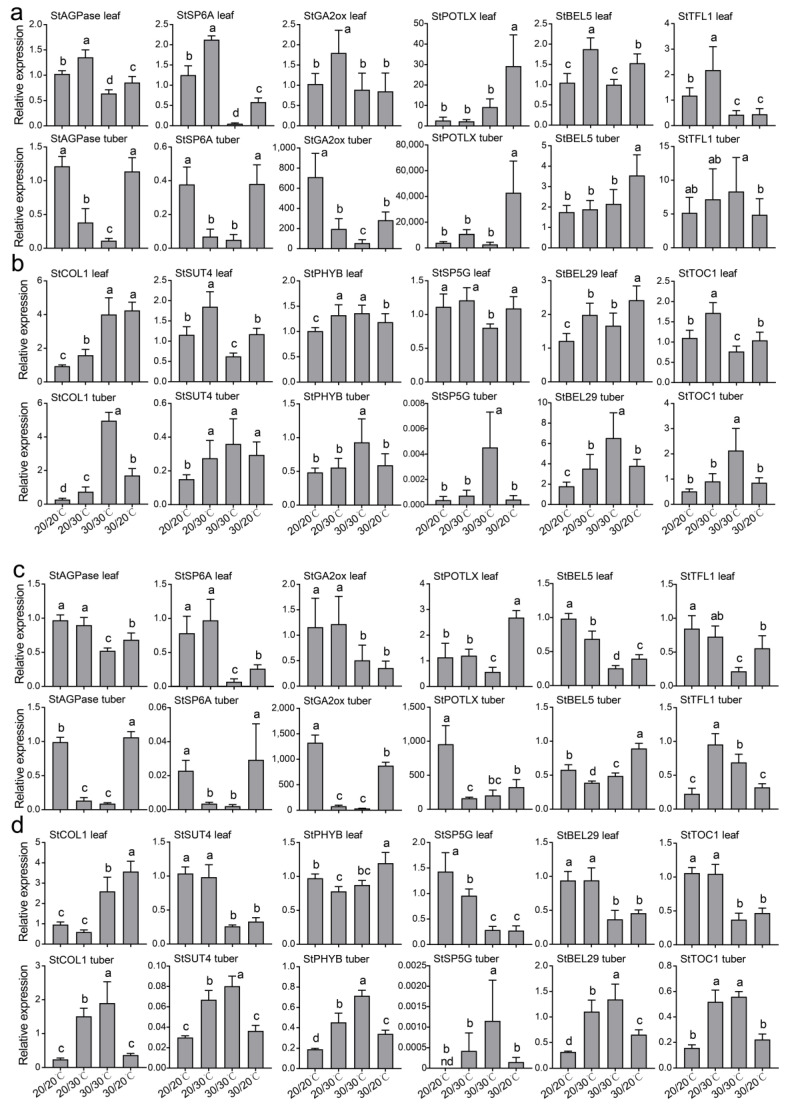
The mechanism of potato (Solanum tuberosum L.) thermotolerance has been the focus of intensive research for many years because plant growth and tuber yield are highly sensitive to heat stress. However, the linkage between the aerial and belowground parts of potato plants in response to high temperatures is not clear. To disentangle this issue, the aerial and belowground parts of the heat-resistant cultivar Dian187 (D187) and the heat-sensitive cultivar Qingshu 9 (Qs9) were independently exposed to high-temperature (30 °C) conditions using a special incubator. The results indicated that when the belowground plant parts were maintained at a normal temperature, the growth of the aerial plant parts was maintained even when independently exposed to heat stress. In contrast, the treatment that independently exposed the belowground plant parts to heat stress promoted premature senescence in the plant's leaves, even when the aerial plant parts were maintained at a normal temperature. When the aerial part of the plant was independently treated with heat stress, tuberization belowground was not delayed, and tuberization suppression was not as severe as when the belowground plant parts independently underwent heat stress. Heat stress on the belowground plant parts alone had virtually no damaging effects on the leaf photosynthetic system but caused distinct tuber deformation, secondary growth, and the loss of tuber skin colour. Transcriptome analysis revealed that the treatment of the belowground plant parts at 30 °C induced 3361 differentially expressed genes in the Qs9 cultivar's expanding tubers, while the D187 cultivar had only 10,148 differentially expressed genes. Conversely, when only the aerial plant parts were treated at 30 °C, there were just 807 DEGs (differentially expressed genes) in the D187 cultivar's expanding tubers compared with 6563 DEGs in the Qs9 cultivar, indicating that the two cultivars with different heat sensitivities have distinct regulatory mechanisms of tuberization when exposed to heat stress. The information provided in this study may be useful for further exploring the genes associated with high-temperature resistance in potato cultivars.
Keywords: different plant parts; heat stress; potato; tuber development.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Pumisutapon P., Topoonyanont N. Moderate-abiotic stress increase in vitro tuberization and microtuber growth of potato. VI Int. Symp. Prod. Establ. Micropropagated Plants. 2015;1155:215–220. doi: 10.17660/ActaHortic.2017.1155.30. - DOI
-
- George T.S., Taylor M.A., Dodd I.C., White P.J. Climate change and consequences for potato production a review of tolerance to emerging abiotic stress. Potato Res. 2017;60:239–268. doi: 10.1007/s11540-018-9366-3. - DOI
-
- Hancock R.D., Morris W.L., Ducreux L.J., Morris J.A., Usman M., Verrall S.R., Fuller J., Simpson C.G., Zhang R.X., Hedley P.E., et al. Physiological biochemical and molecular responses of the potato (Solanum tuberosum L.) plant to moderately elevated temperature. Plant Cell Environ. 2014;37:439–450. doi: 10.1111/pce.12168. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
