

A density functional theory for ecology across scales
- PMID: 36841818
- PMCID: PMC9968302
- DOI: 10.1038/s41467-023-36628-4
A density functional theory for ecology across scales
Abstract
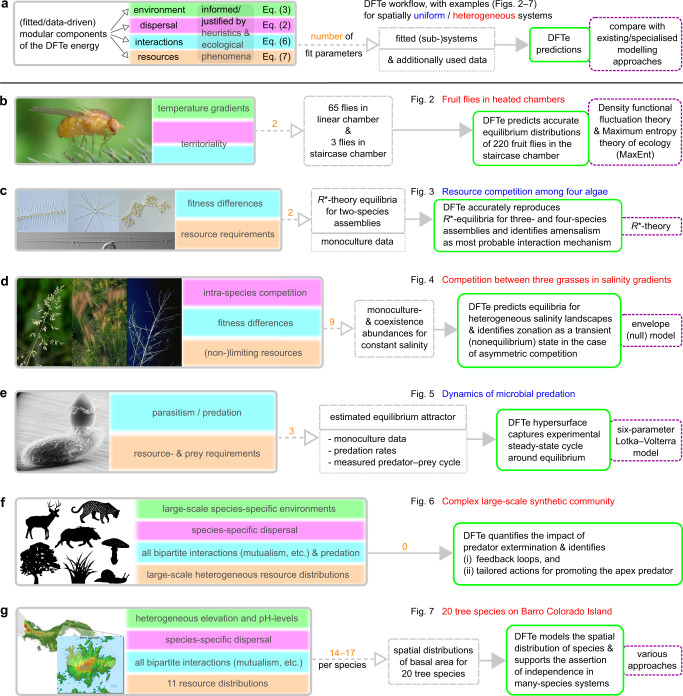
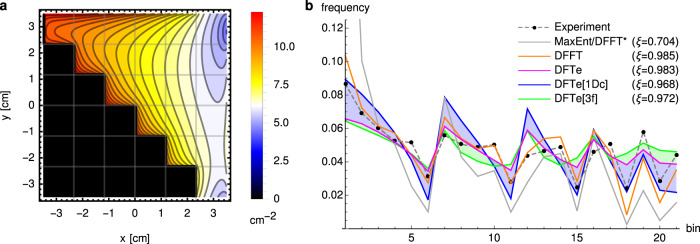
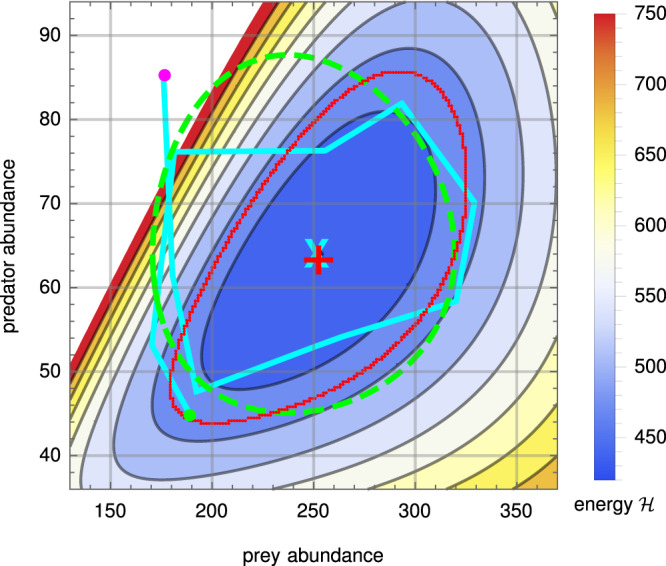
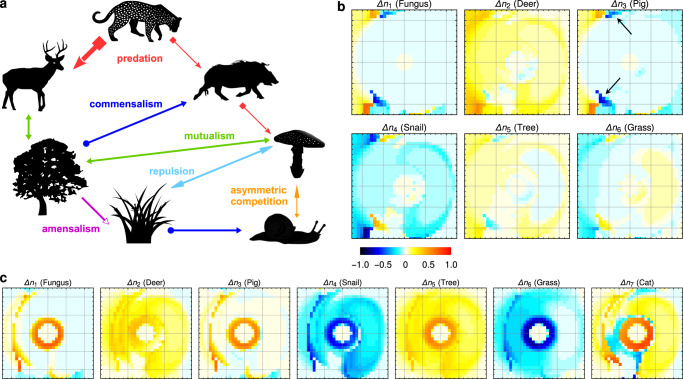
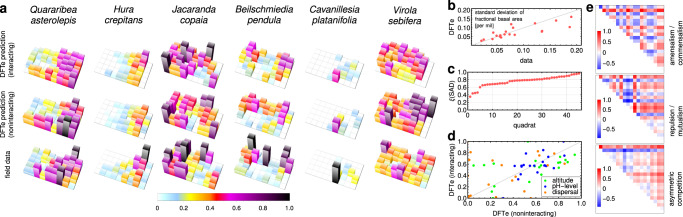
Ecology lacks a holistic approach that can model phenomena across temporal and spatial scales, largely because of the challenges in modelling systems with a large number of interacting constituents. This hampers our understanding of complex ecosystems and the impact that human interventions (e.g., deforestation, wildlife harvesting and climate change) have on them. Here we use density functional theory, a computational method for many-body problems in physics, to develop a computational framework for ecosystem modelling. Our methods accurately fit experimental and synthetic data of interacting multi-species communities across spatial scales and can project to unseen data. As the key concept we establish and validate a cost function that encodes the trade-offs between the various ecosystem components. We show how this single general modelling framework delivers predictions on par with established, but specialised, approaches for systems from predatory microbes to territorial flies to tropical tree communities. Our density functional framework thus provides a promising avenue for advancing our understanding of ecological systems.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
The ecology, distribution, conservation and management of large old trees.Biol Rev Camb Philos Soc. 2017 Aug;92(3):1434-1458. doi: 10.1111/brv.12290. Epub 2016 Jul 7. Biol Rev Camb Philos Soc. 2017. PMID: 27383287 Review.
-
Spatial Distribution Patterns in the Very Rare and Species-Rich Picea chihuahuana Tree Community (Mexico).PLoS One. 2015 Oct 23;10(10):e0140442. doi: 10.1371/journal.pone.0140442. eCollection 2015. PLoS One. 2015. PMID: 26496189 Free PMC article.
-
Intra- and interspecific tree growth across a long altitudinal gradient in the Peruvian Andes.Ecology. 2012 Sep;93(9):2061-72. doi: 10.1890/11-1725.1. Ecology. 2012. PMID: 23094378
-
A guide to ecosystem models and their environmental applications.Nat Ecol Evol. 2020 Nov;4(11):1459-1471. doi: 10.1038/s41559-020-01298-8. Epub 2020 Sep 14. Nat Ecol Evol. 2020. PMID: 32929239 Review.
-
Woody-plant ecosystems under climate change and air pollution-response consistencies across zonobiomes?Tree Physiol. 2017 Jun 1;37(6):706-732. doi: 10.1093/treephys/tpx009. Tree Physiol. 2017. PMID: 28338970
Cited by
-
Assessments of various precipitation product performances and disaster monitoring utilities over the Tibetan Plateau.Sci Rep. 2024 Aug 26;14(1):19740. doi: 10.1038/s41598-024-70547-8. Sci Rep. 2024. PMID: 39187512 Free PMC article.
References
-
- Tilman D. Tests of resource competition theory using four species of Lake Michigan Algae. Ecology. 1981;62:802. doi: 10.2307/1937747. - DOI
-
- Pacala SW, et al. Forest models defined by field measurements: estimation, error analysis and dynamics. Ecol. Monogr. 1996;66:1. doi: 10.2307/2963479. - DOI
-
- Cabral JS, Valente L, Hartig F. Mechanistic simulation models in macroecology and biogeography: state-of-art and prospects. Ecography. 2017;40:267. doi: 10.1111/ecog.02480. - DOI
-
- Levin SA. The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology. 1992;73:1943. doi: 10.2307/1941447. - DOI
-
- Leibold MA, et al. The metacommunity concept: a framework for multi-scale community ecology. Ecol. Lett. 2004;7:601. doi: 10.1111/j.1461-0248.2004.00608.x. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
