

Stress-induced plasticity of a CRH/GABA projection disrupts reward behaviors in mice
- PMID: 36841826
- PMCID: PMC9968307
- DOI: 10.1038/s41467-023-36780-x
Stress-induced plasticity of a CRH/GABA projection disrupts reward behaviors in mice
Abstract
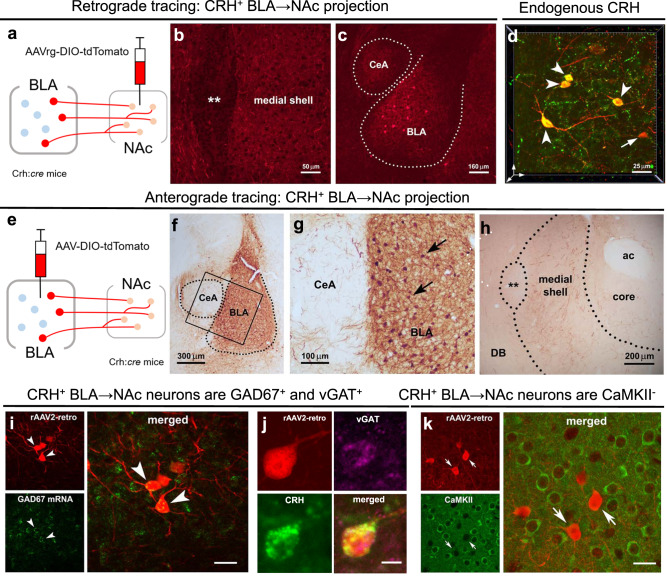
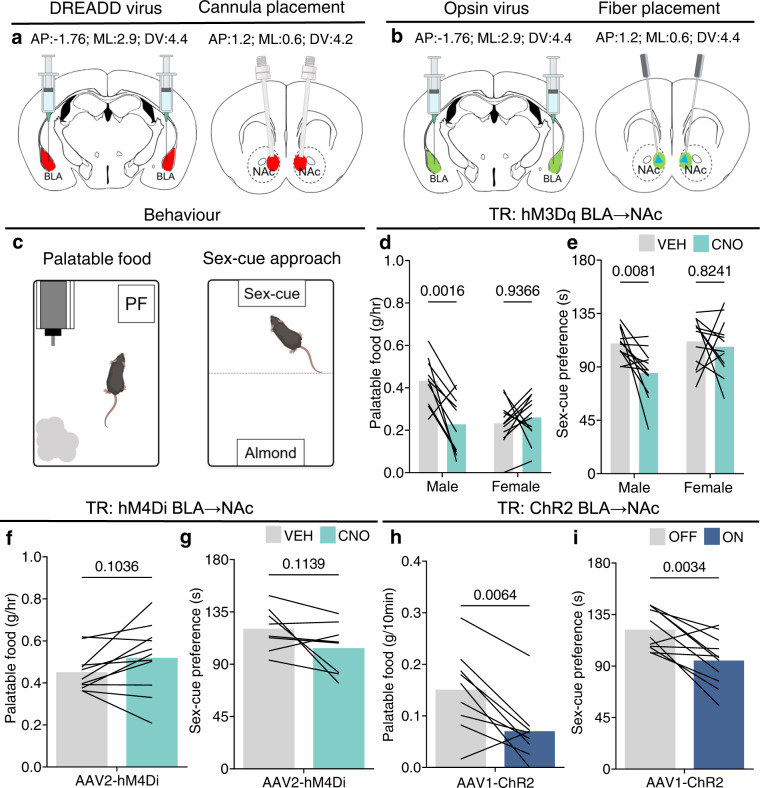
Disrupted operations of the reward circuit underlie major emotional disorders, including depression, which commonly arise following early life stress / adversity (ELA). However, how ELA enduringly impacts reward circuit functions remains unclear. We characterize a stress-sensitive projection connecting basolateral amygdala (BLA) and nucleus accumbens (NAc) that co-expresses GABA and the stress-reactive neuropeptide corticotropin-releasing hormone (CRH). We identify a crucial role for this projection in executing disrupted reward behaviors provoked by ELA: chemogenetic and optogenetic stimulation of the projection in control male mice suppresses several reward behaviors, recapitulating deficits resulting from ELA and demonstrating the pathway's contributions to normal reward behaviors. In adult ELA mice, inhibiting-but not stimulating-the projection, restores typical reward behaviors yet has little effect in controls, indicating ELA-induced maladaptive plasticity of this reward-circuit component. Thus, we discover a stress-sensitive, reward inhibiting BLA → NAc projection with unique molecular features, which may provide intervention targets for disabling mental illnesses.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
