

The Colorectal Cancer Microbiota Alter Their Transcriptome To Adapt to the Acidity, Reactive Oxygen Species, and Metabolite Availability of Gut Microenvironments
- PMID: 36847536
- PMCID: PMC10117117
- DOI: 10.1128/msphere.00627-22
The Colorectal Cancer Microbiota Alter Their Transcriptome To Adapt to the Acidity, Reactive Oxygen Species, and Metabolite Availability of Gut Microenvironments
Abstract
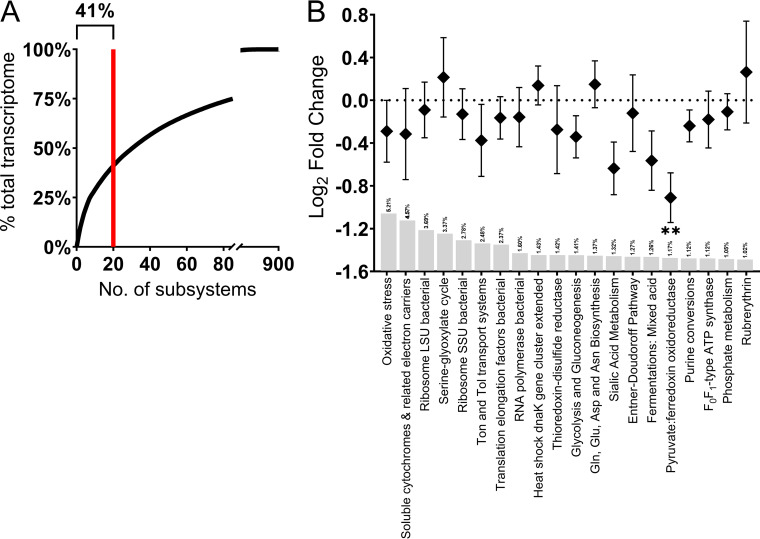
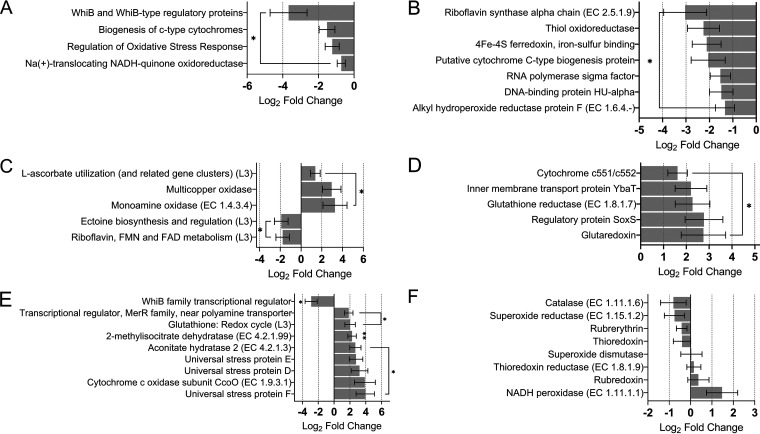
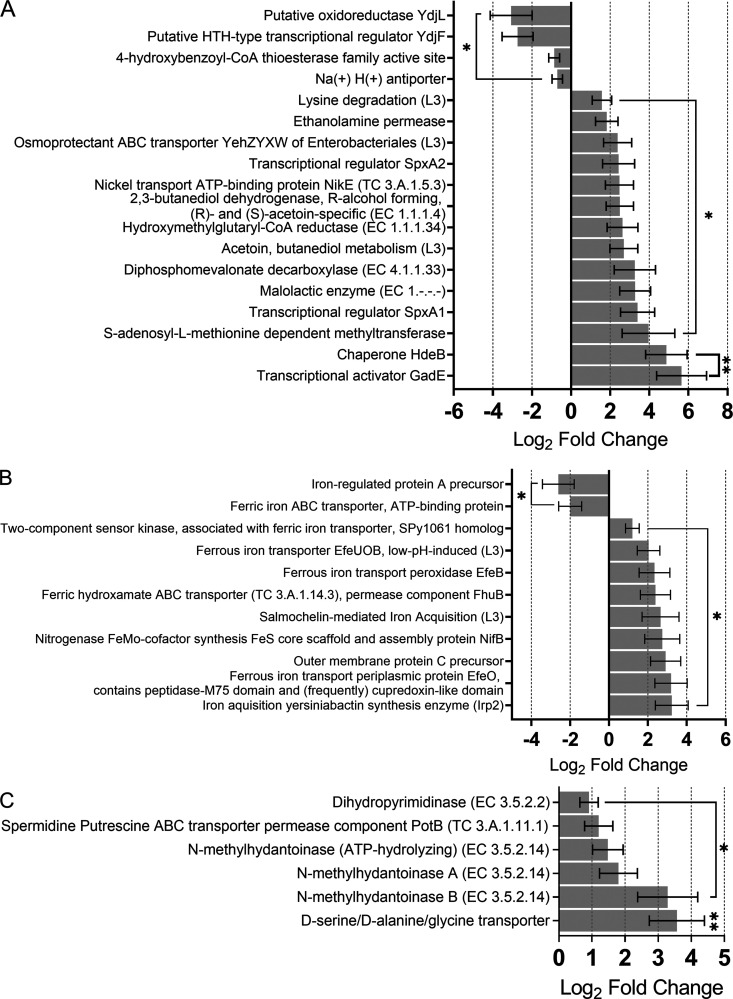
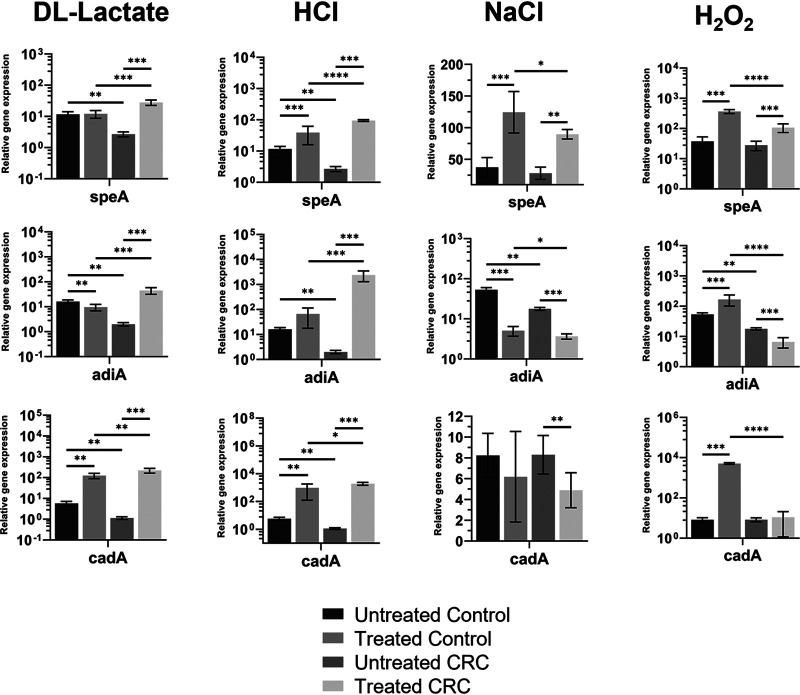
The gut microbiome is implicated in the pathology of colorectal cancer (CRC). However, the mechanisms by which the microbiota actively contribute to disease onset and progression remain elusive. In this pilot study, we sequenced fecal metatranscriptomes of 10 non-CRC and 10 CRC patient gut microbiomes and conducted differential gene expression analyses to assess any changed functionality in disease. We report that oxidative stress responses were the dominant activity across cohorts, an overlooked protective housekeeping role of the human gut microbiome. However, expression of hydrogen peroxide and nitric oxide-scavenging genes was diminished and augmented, respectively, positing that these regulated microbial responses have implications for CRC pathology. CRC microbes enhanced expression of genes for host colonization, biofilm formation, genetic exchange, virulence determinants, antibiotic, and acid resistances. Moreover, microbes promoted transcription of genes involved in metabolism of several beneficial metabolites, suggesting their contribution to patient metabolite deficiencies previously solely attributed to tumor cells. We showed in vitro that expression of genes involved in amino acid-dependent acid resistance mechanisms of meta-gut Escherichia coli responded differently to acid, salt, and oxidative pressures under aerobic conditions. These responses were mostly dictated by the host health status of origin of the microbiota, suggesting their exposure to fundamentally different gut conditions. These findings for the first time highlight mechanisms by which the gut microbiota can either protect against or drive colorectal cancer and provide insights into the cancerous gut environment that drives functional characteristics of the microbiome. IMPORTANCE The human gut microbiota has the genetic potential to drive colorectal cancer onset and progression; however, the expression of this genetic potential during the disease has not been investigated. We found that microbial expression of genes that detoxify DNA-damaging reactive oxygen species, which drive colorectal cancer, is compromised in cancer. We observed a greater activation of expression of genes involved in virulence, host colonization, exchange of genetic material, metabolite utilization, defense against antibiotics, and environmental pressures. Culturing gut Escherichia coli of cancerous and noncancerous metamicrobiota revealed different regulatory responses of amino acid-dependent acid resistance mechanisms in a health-dependent manner under environmental acid, oxidative, and osmotic pressures. Here, for the first time, we demonstrate that the activity of microbial genomes is regulated by the health status of the gut in vivo and in vitro and provides new insights for shifts in microbial gene expression in colorectal cancer.
Keywords: acidity; colorectal cancer; gut microbiota; metatranscriptome; reactive oxygen species; virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
The Colorectal Cancer Gut Environment Regulates Activity of the Microbiome and Promotes the Multidrug Resistant Phenotype of ESKAPE and Other Pathogens.mSphere. 2023 Apr 20;8(2):e0062622. doi: 10.1128/msphere.00626-22. Epub 2023 Feb 27. mSphere. 2023. PMID: 36847529 Free PMC article.
-
Association analysis of gut microbiota with LDL-C metabolism and microbial pathogenicity in colorectal cancer patients.Lipids Health Dis. 2024 Nov 8;23(1):367. doi: 10.1186/s12944-024-02333-4. Lipids Health Dis. 2024. PMID: 39516755 Free PMC article.
-
Inhibition of the NF-κB/HIF-1α signaling pathway in colorectal cancer by tyrosol: a gut microbiota-derived metabolite.J Immunother Cancer. 2024 Sep 28;12(9):e008831. doi: 10.1136/jitc-2024-008831. J Immunother Cancer. 2024. PMID: 39343509 Free PMC article.
-
Gut Microbiota and Colorectal Cancer: A Balance Between Risk and Protection.Int J Mol Sci. 2025 Apr 15;26(8):3733. doi: 10.3390/ijms26083733. Int J Mol Sci. 2025. PMID: 40332367 Free PMC article. Review.
-
Role of the Gut Microbiota and Its Metabolites in Tumorigenesis or Development of Colorectal Cancer.Adv Sci (Weinh). 2023 Aug;10(23):e2205563. doi: 10.1002/advs.202205563. Epub 2023 Jun 1. Adv Sci (Weinh). 2023. PMID: 37263983 Free PMC article. Review.
Cited by
-
Oxidative Stress and Age-Related Tumors.Antioxidants (Basel). 2024 Sep 13;13(9):1109. doi: 10.3390/antiox13091109. Antioxidants (Basel). 2024. PMID: 39334768 Free PMC article. Review.
-
Gut metatranscriptomics based de novo assembly reveals microbial signatures predicting immunotherapy outcomes in non-small cell lung cancer.J Transl Med. 2024 Nov 19;22(1):1044. doi: 10.1186/s12967-024-05835-y. J Transl Med. 2024. PMID: 39563352 Free PMC article.
-
The Colorectal Cancer Gut Environment Regulates Activity of the Microbiome and Promotes the Multidrug Resistant Phenotype of ESKAPE and Other Pathogens.mSphere. 2023 Apr 20;8(2):e0062622. doi: 10.1128/msphere.00626-22. Epub 2023 Feb 27. mSphere. 2023. PMID: 36847529 Free PMC article.
-
Fusobacterium species are distinctly associated with patients with Lynch syndrome colorectal cancer.iScience. 2024 Jun 4;27(7):110181. doi: 10.1016/j.isci.2024.110181. eCollection 2024 Jul 19. iScience. 2024. PMID: 38993678 Free PMC article.
-
In Vitro Influence of Specific Bacteroidales Strains on Gut and Liver Health Related to Metabolic Dysfunction-Associated Fatty Liver Disease.Probiotics Antimicrob Proteins. 2025 Jun;17(3):1498-1512. doi: 10.1007/s12602-024-10219-1. Epub 2024 Feb 6. Probiotics Antimicrob Proteins. 2025. PMID: 38319537 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
