

Comparative and Functional Analyses Reveal Conserved and Variable Regulatory Systems That Control Lasalocid Biosynthesis in Different Streptomyces Species
- PMID: 36847561
- PMCID: PMC10100954
- DOI: 10.1128/spectrum.03852-22
Comparative and Functional Analyses Reveal Conserved and Variable Regulatory Systems That Control Lasalocid Biosynthesis in Different Streptomyces Species
Abstract
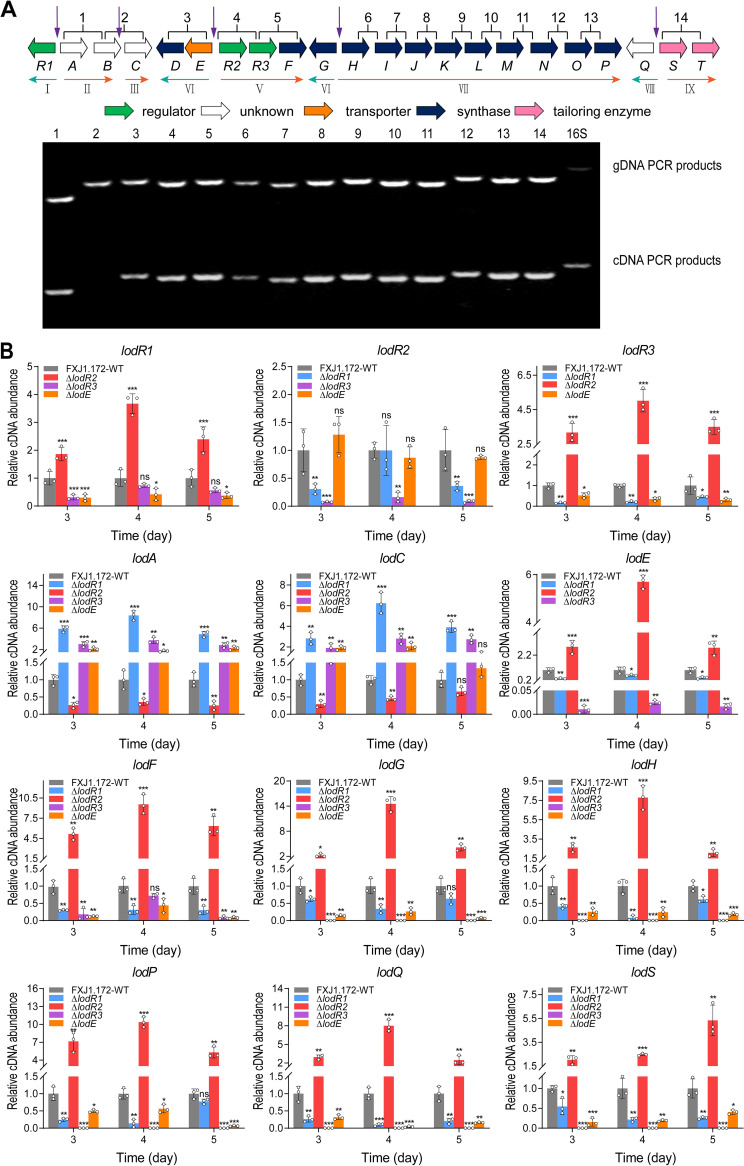
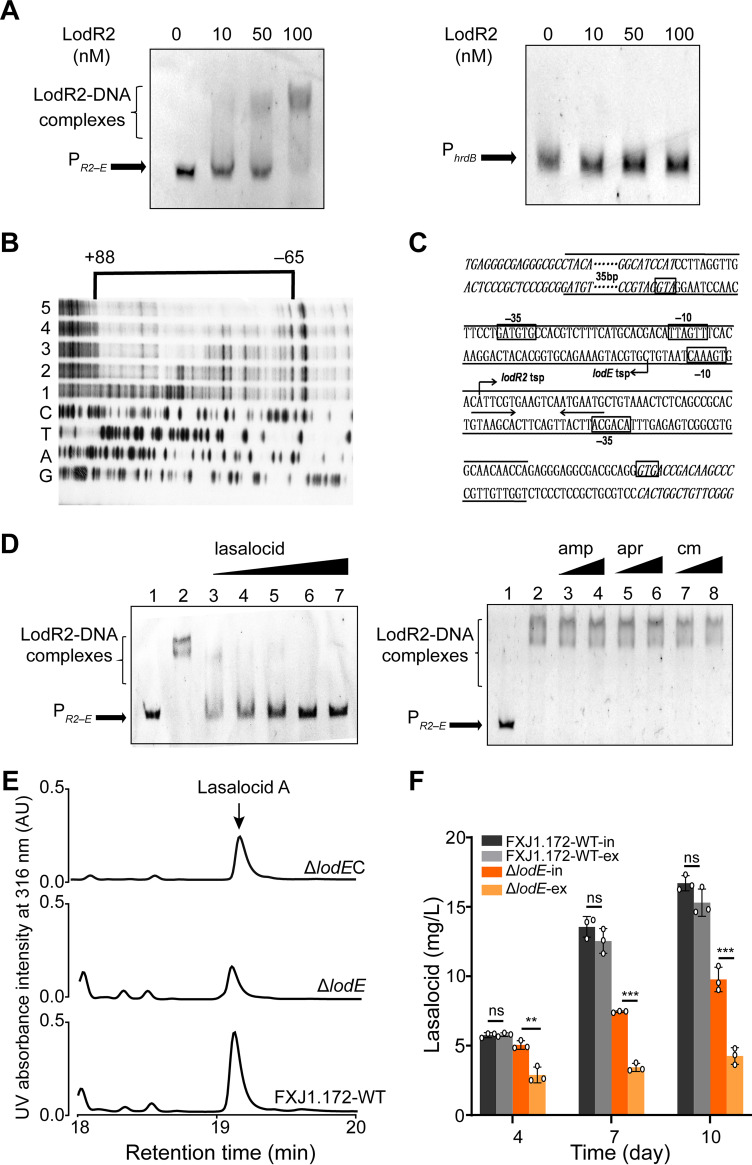
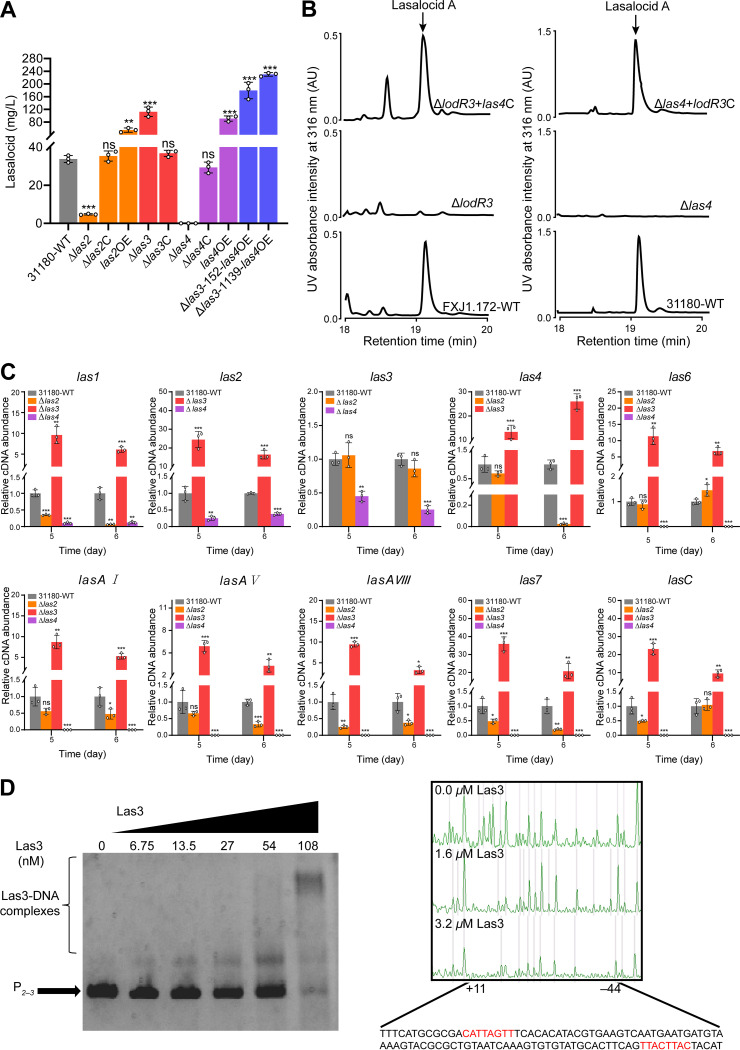
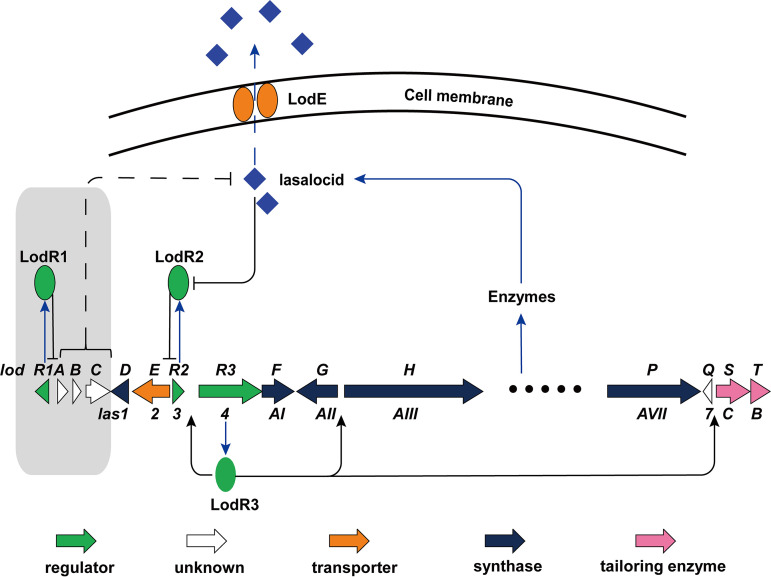
Lasalocid, a representative polyether ionophore, has been successfully applied in veterinary medicine and animal husbandry and also displays promising potential for cancer therapy. Nevertheless, the regulatory system governing lasalocid biosynthesis remains obscure. Here, we identified two conserved (lodR2 and lodR3) and one variable (lodR1, found only in Streptomyces sp. strain FXJ1.172) putative regulatory genes through a comparison of the lasalocid biosynthetic gene cluster (lod) from Streptomyces sp. FXJ1.172 with those (las and lsd) from Streptomyces lasalocidi. Gene disruption experiments demonstrated that both lodR1 and lodR3 positively regulate lasalocid biosynthesis in Streptomyces sp. FXJ1.172, while lodR2 plays a negative regulatory role. To unravel the regulatory mechanism, transcriptional analysis and electrophoretic mobility shift assays (EMSAs) along with footprinting experiments were performed. The results revealed that LodR1 and LodR2 could bind to the intergenic regions of lodR1-lodAB and lodR2-lodED, respectively, thereby repressing the transcription of the lodAB and lodED operons, respectively. The repression of lodAB-lodC by LodR1 likely boosts lasalocid biosynthesis. Furthermore, LodR2 and LodE constitute a repressor-activator system that senses changes in intracellular lasalocid concentrations and coordinates its biosynthesis. LodR3 could directly activate the transcription of key structural genes. Comparative and parallel functional analyses of the homologous genes in S. lasalocidi ATCC 31180T confirmed the conserved roles of lodR2, lodE, and lodR3 in controlling lasalocid biosynthesis. Intriguingly, the variable gene locus lodR1-lodC from Streptomyces sp. FXJ1.172 seems functionally conserved when introduced into S. lasalocidi ATCC 31180T. Overall, our findings demonstrate that lasalocid biosynthesis is tightly controlled by both conserved and variable regulators, providing valuable guidance for further improving lasalocid production. IMPORTANCE Compared to its elaborated biosynthetic pathway, the regulation of lasalocid biosynthesis remains obscure. Here, we characterize the roles of regulatory genes in lasalocid biosynthetic gene clusters of two distinct Streptomyces species and identify a conserved repressor-activator system, LodR2-LodE, which could sense changes in the concentration of lasalocid and coordinate its biosynthesis with self-resistance. Furthermore, in parallel, we verify that the regulatory system identified in a new Streptomyces isolate is valid in the industrial lasalocid producer and thus applicable for the construction of high-yield strains. These findings deepen our understanding of regulatory mechanisms involved in the production of polyether ionophores and provide novel clues for the rational design of industrial strains for scaled-up production.
Keywords: Streptomyces; biosynthesis regulation; biosynthetic gene cluster; lasalocid; regulatory diversification; repressor-activator system.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Mitrovic M, Schildknecht EG, Marusich WL. 1975. Comparative anticoccidial activity and compatibility of lasalocid in broiler chickens. Poult Sci 54:757–761. doi:10.3382/ps.0540757. - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
