

Constitutive and induced forms of membrane-bound proteinase 3 interact with antineutrophil cytoplasmic antibodies and promote immune activation of neutrophils
- PMID: 36849007
- PMCID: PMC10124916
- DOI: 10.1016/j.jbc.2023.103072
Constitutive and induced forms of membrane-bound proteinase 3 interact with antineutrophil cytoplasmic antibodies and promote immune activation of neutrophils
Abstract
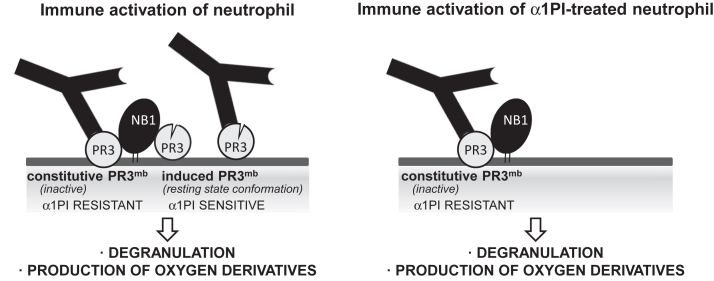
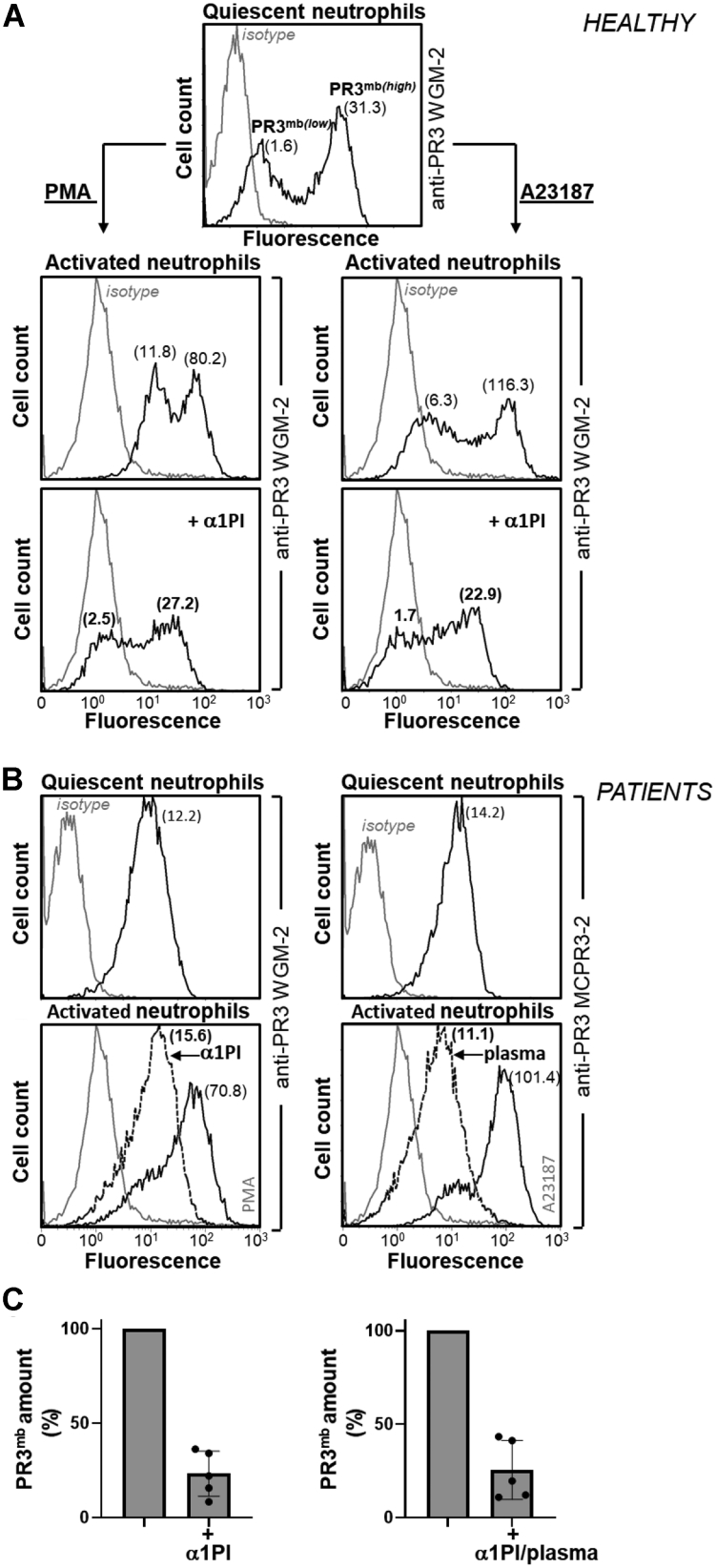
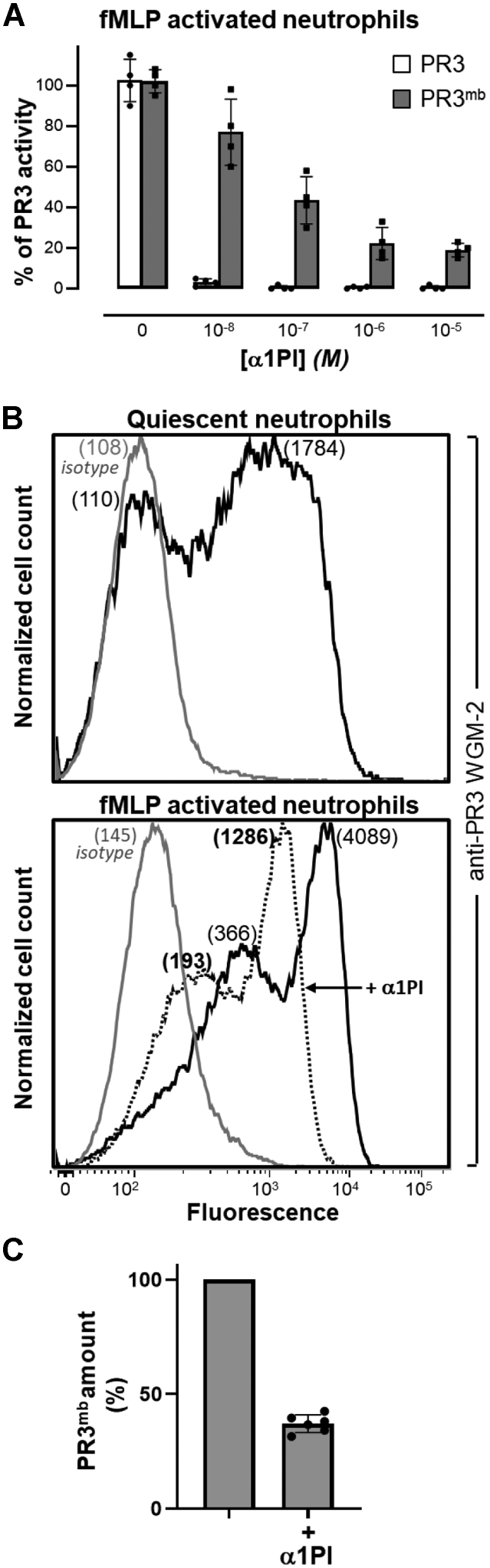
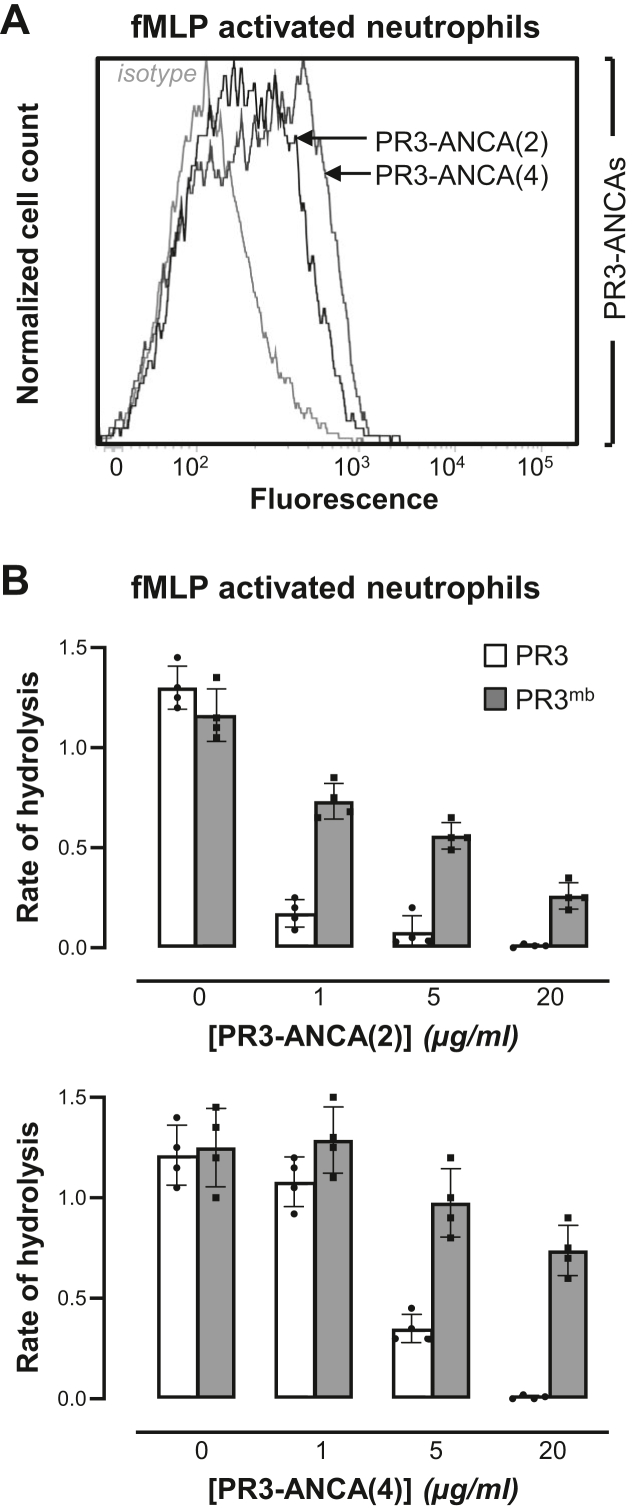
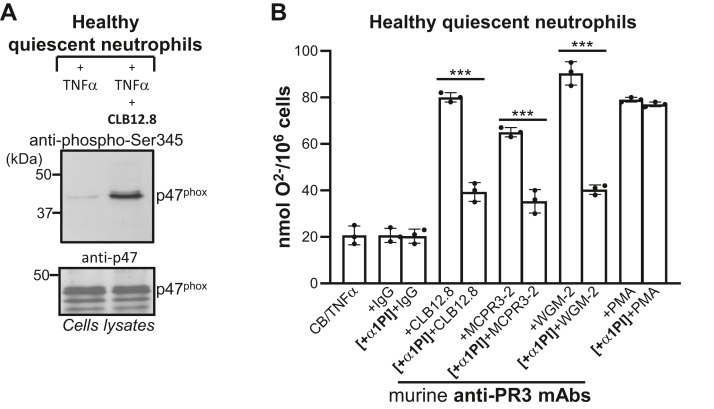
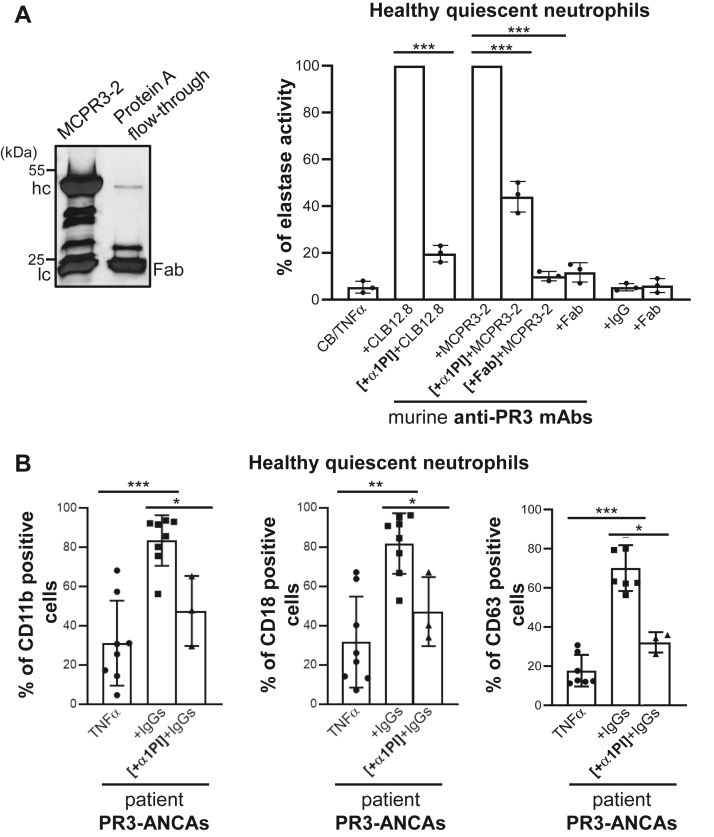
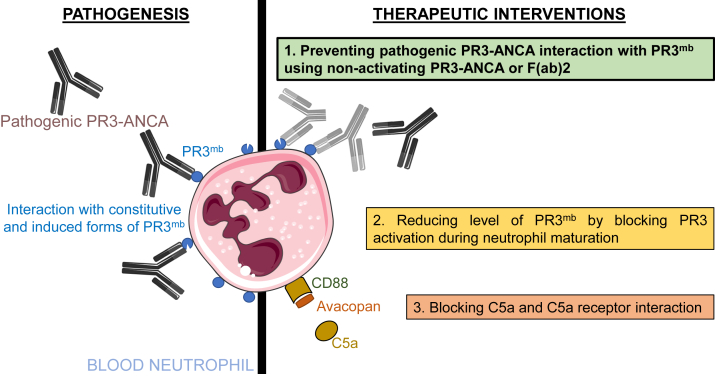
Proteinase 3 (PR3) is the main target antigen of antineutrophil cytoplasmic antibodies (ANCAs) in PR3-ANCA-associated vasculitis. A small fraction of PR3 is constitutively exposed on the surface of quiescent blood neutrophils in a proteolytically inactive form. When activated, neutrophils expose an induced form of membrane-bound PR3 (PR3mb) on their surface as well, which is enzymatically less active than unbound PR3 in solution due to its altered conformation. In this work, our objective was to understand the respective role of constitutive and induced PR3mb in the immune activation of neutrophils triggered by murine anti-PR3 mAbs and human PR3-ANCA. We quantified immune activation of neutrophils by the measurement of the production of superoxide anions and secreted protease activity in the cell supernatant before and after treatment of the cells by alpha-1 protease inhibitor that clears induced PR3mb from the cell surface. Incubation of TNFα-primed neutrophils with anti-PR3 antibodies resulted in a significant increase in superoxide anion production, membrane activation marker exposition, and secreted protease activity. When primed neutrophils were first treated with alpha-1 protease inhibitor, we observed a partial reduction in antibody-induced neutrophil activation, suggesting that constitutive PR3mb is sufficient to activate neutrophils. The pretreatment of primed neutrophils with purified antigen-binding fragments used as competitor significantly reduced cell activation by whole antibodies. This led us to the conclusion that PR3mb promoted immune activation of neutrophils. We propose that blocking and/or elimination of PR3mb offers a new therapeutic strategy to attenuate neutrophil activation in patients with PR3-ANCA-associated vasculitis.
Keywords: PR3-ANCA; alpha-1 protease inhibitor; anti-PR3 mAbs; autoantibody; immune activation; inflammation; neutrophil; proteinase 3; therapeutic strategy; vasculitis.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Kitching A.R., Anders H.J., Basu N., Brouwer E., Gordon J., Jayne D.R., et al. ANCA-associated vasculitis. Nat. Rev. Dis. Primers. 2020;6:71. - PubMed
-
- Kettritz R. Vasculitis: a clear argument for targeting complement in ANCA vasculitis. Nat. Rev. Nephrol. 2017;13:448–450. - PubMed
-
- Korkmaz B., Lesner A., Guarino C., Wysocka M., Kellenberger C., Watier H., et al. Inhibitors and antibody fragments as potential anti-inflammatory therapeutics targeting neutrophil proteinase 3 in human disease. Pharmacol. Rev. 2016;68:603–630. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
