

Structural modelling of human complement FHR1 and two of its synthetic derivatives provides insight into their in-vivo functions
- PMID: 36851916
- PMCID: PMC9957715
- DOI: 10.1016/j.csbj.2023.02.002
Structural modelling of human complement FHR1 and two of its synthetic derivatives provides insight into their in-vivo functions
Abstract
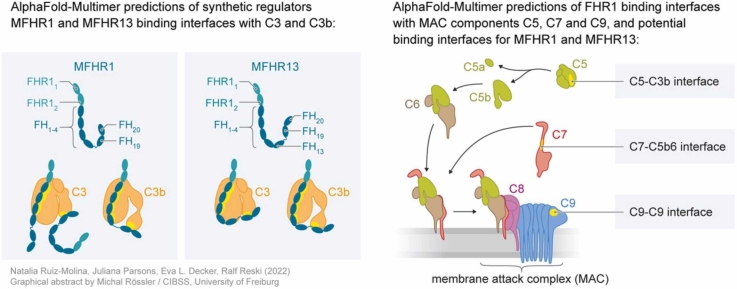
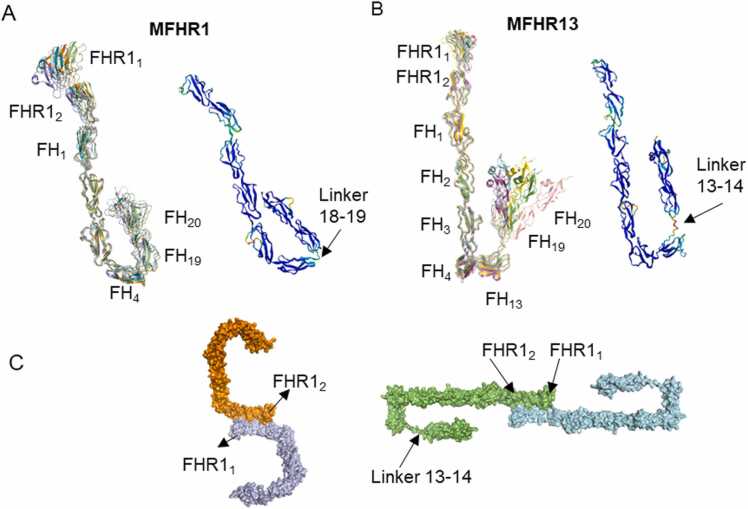
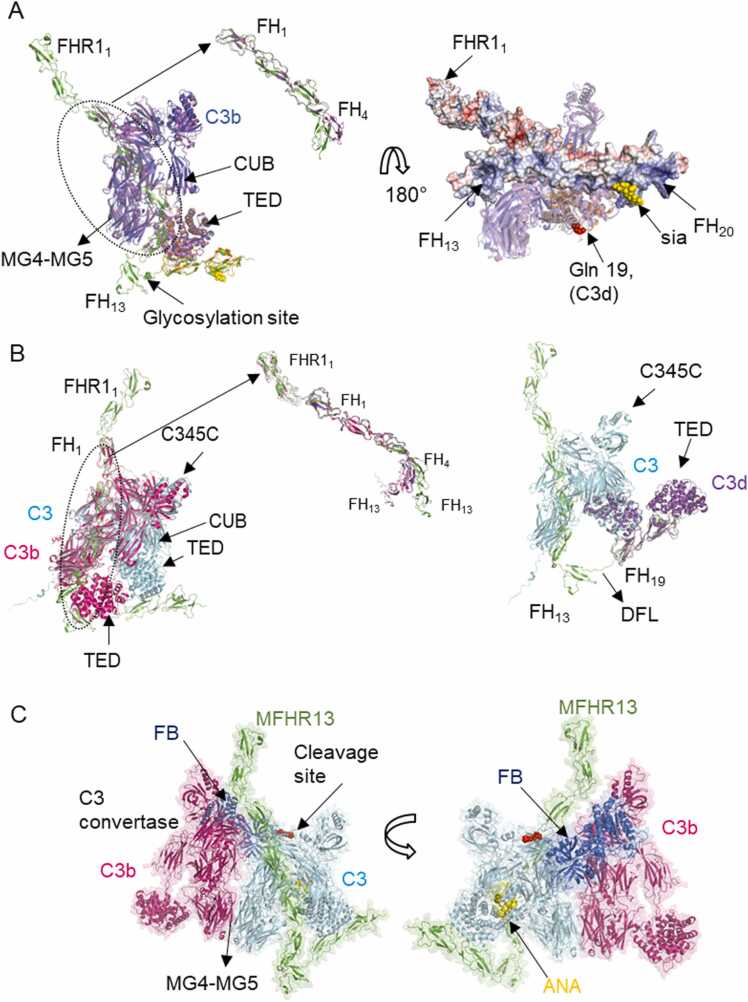
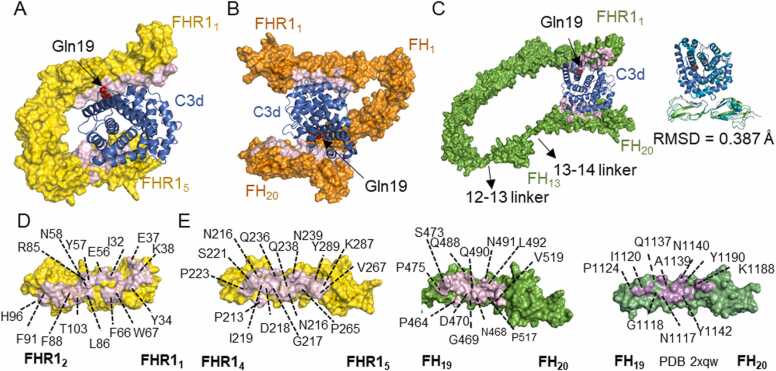
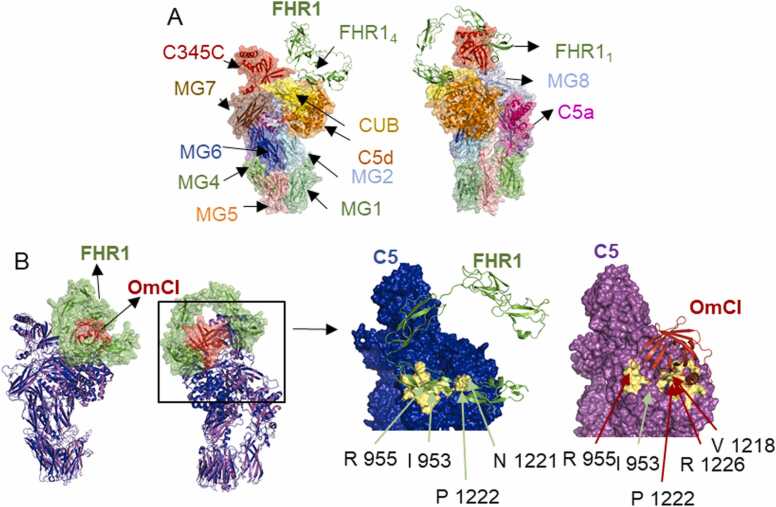
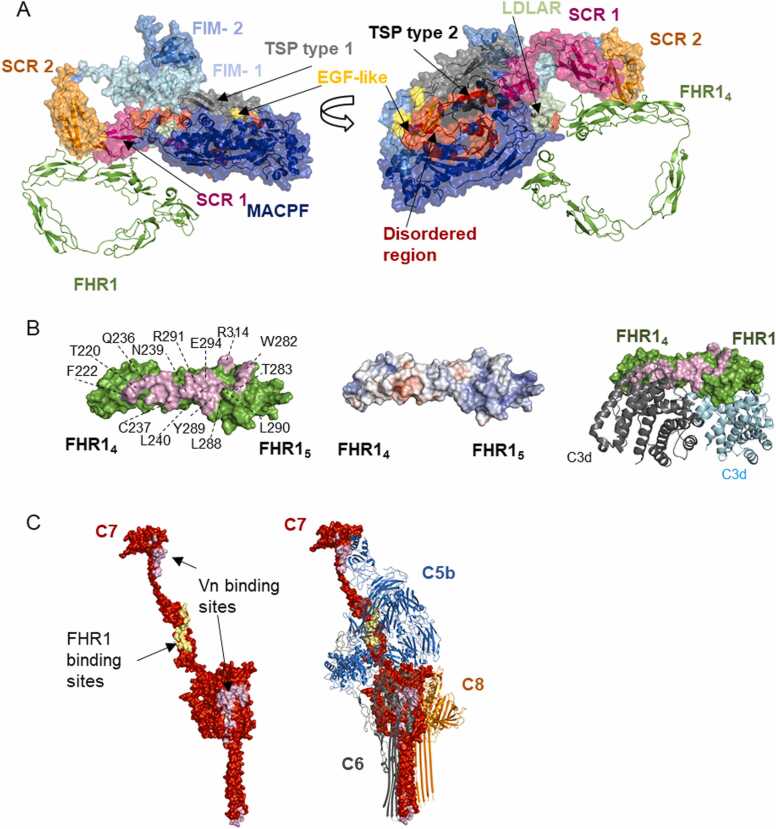
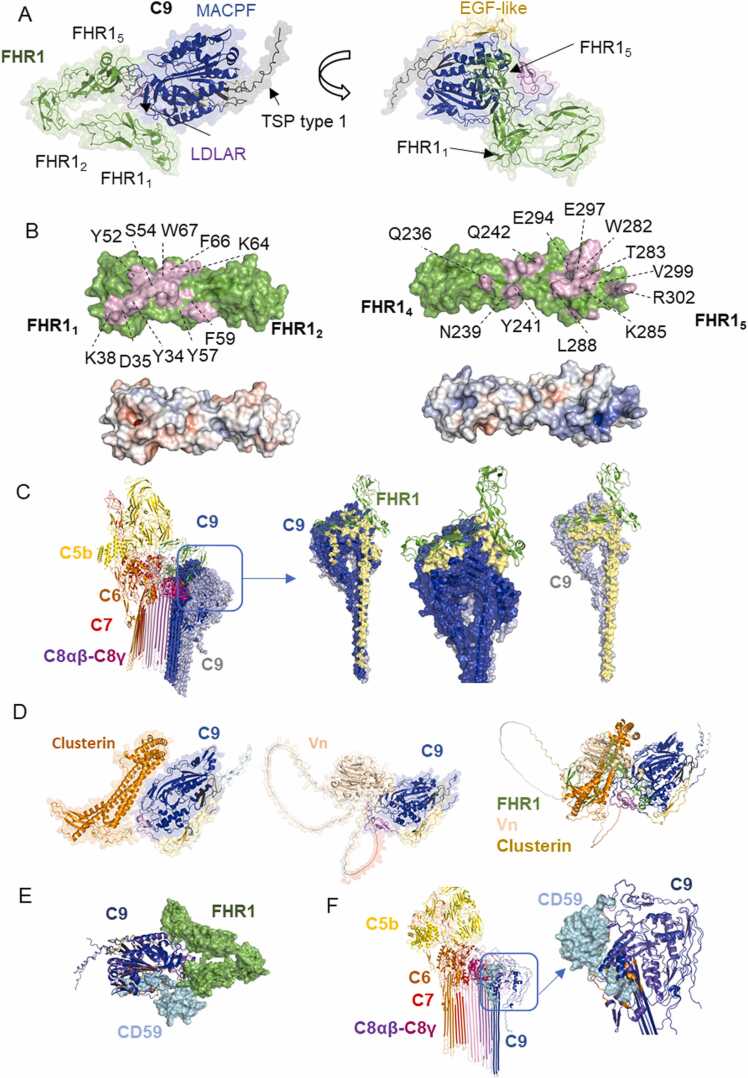
Human complement is the first line of defence against invading pathogens and is involved in tissue homeostasis. Complement-targeted therapies to treat several diseases caused by a dysregulated complement are highly desirable. Despite huge efforts invested in their development, only very few are currently available, and a deeper understanding of the numerous interactions and complement regulation mechanisms is indispensable. Two important complement regulators are human Factor H (FH) and Factor H-related protein 1 (FHR1). MFHR1 and MFHR13, two promising therapeutic candidates based on these regulators, combine the dimerization and C5-regulatory domains of FHR1 with the central C3-regulatory and cell surface-recognition domains of FH. Here, we used AlphaFold2 to model the structure of these two synthetic regulators. Moreover, we used AlphaFold-Multimer (AFM) to study possible interactions of C3 fragments and membrane attack complex (MAC) components C5, C7 and C9 in complex with FHR1, MFHR1, MFHR13 as well as the best-known MAC regulators vitronectin (Vn), clusterin and CD59, whose experimental structures remain undetermined. AFM successfully predicted the binding interfaces of FHR1 and the synthetic regulators with C3 fragments and suggested binding to C3. The models revealed structural differences in binding to these ligands through different interfaces. Additionally, AFM predictions of Vn, clusterin or CD59 with C7 or C9 agreed with previously published experimental results. Because the role of FHR1 as MAC regulator has been controversial, we analysed possible interactions with C5, C7 and C9. AFM predicted interactions of FHR1 with proteins of the terminal complement complex (TCC) as indicated by experimental observations, and located the interfaces in FHR11-2 and FHR14-5. According to AFM prediction, FHR1 might partially block the C3b binding site in C5, inhibiting C5 activation, and block C5b-7 complex formation and C9 polymerization, with similar mechanisms of action as clusterin and vitronectin. Here, we generate hypotheses and give the basis for the design of rational approaches to understand the molecular mechanism of MAC inhibition, which will facilitate the development of further complement therapeutics.
Keywords: AlphaFold; Complement factor H-related 1; Complement regulation; Complement therapeutics; Factor H; Membrane attack complex.
© 2023 The Authors.
Conflict of interest statement
All authors declare to have no competing interests.
Figures
Similar articles
-
Complement Factor H-Related Proteins FHR1 and FHR5 Interact With Extracellular Matrix Ligands, Reduce Factor H Regulatory Activity and Enhance Complement Activation.Front Immunol. 2022 Mar 22;13:845953. doi: 10.3389/fimmu.2022.845953. eCollection 2022. Front Immunol. 2022. PMID: 35392081 Free PMC article.
-
A synthetic protein as efficient multitarget regulator against complement over-activation.Commun Biol. 2022 Feb 22;5(1):152. doi: 10.1038/s42003-022-03094-5. Commun Biol. 2022. PMID: 35194132 Free PMC article.
-
The MFHR1 Fusion Protein Is a Novel Synthetic Multitarget Complement Inhibitor with Therapeutic Potential.J Am Soc Nephrol. 2018 Apr;29(4):1141-1153. doi: 10.1681/ASN.2017070738. Epub 2018 Jan 15. J Am Soc Nephrol. 2018. PMID: 29335241 Free PMC article.
-
The killer molecule of complement.J Invest Dermatol. 1985 Jul;85(1 Suppl):47s-52s. doi: 10.1111/1523-1747.ep12275445. J Invest Dermatol. 1985. PMID: 3891882 Review.
-
Transmembrane channel-formation by five complement proteins.Biochem Soc Symp. 1985;50:235-46. Biochem Soc Symp. 1985. PMID: 2428370 Review.
Cited by
-
Medical Relevance, State-of-the-Art and Perspectives of "Sweet Metacode" in Liquid Biopsy Approaches.Diagnostics (Basel). 2024 Mar 28;14(7):713. doi: 10.3390/diagnostics14070713. Diagnostics (Basel). 2024. PMID: 38611626 Free PMC article. Review.
-
Plasma proteome fingerprint in kidney diseases.Front Mol Biosci. 2025 Jan 17;11:1494779. doi: 10.3389/fmolb.2024.1494779. eCollection 2024. Front Mol Biosci. 2025. PMID: 39896931 Free PMC article.
-
Differential prolyl hydroxylation by six Physcomitrella prolyl-4 hydroxylases.Comput Struct Biotechnol J. 2024 Jun 13;23:2580-2594. doi: 10.1016/j.csbj.2024.06.014. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 39021582 Free PMC article.
References
-
- Zipfel P.F., Skerka C. Complement regulators and inhibitory proteins. Nat Rev Immunol. 2009;9:729–740. - PubMed
-
- Mannes M., Dopler A., Zolk O., Lang S.J., Halbgebauer R., Höchsmann B., et al. Complement inhibition at the level of C3 or C5: Mechanistic reasons for ongoing terminal pathway activity. Blood. 2021;137:443–455. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous