

An evolutionarily conserved bimodular domain anchors ZC3HC1 and its yeast homologue Pml39p to the nuclear basket
- PMID: 36857168
- PMCID: PMC10162418
- DOI: 10.1091/mbc.E22-09-0402
An evolutionarily conserved bimodular domain anchors ZC3HC1 and its yeast homologue Pml39p to the nuclear basket
Abstract
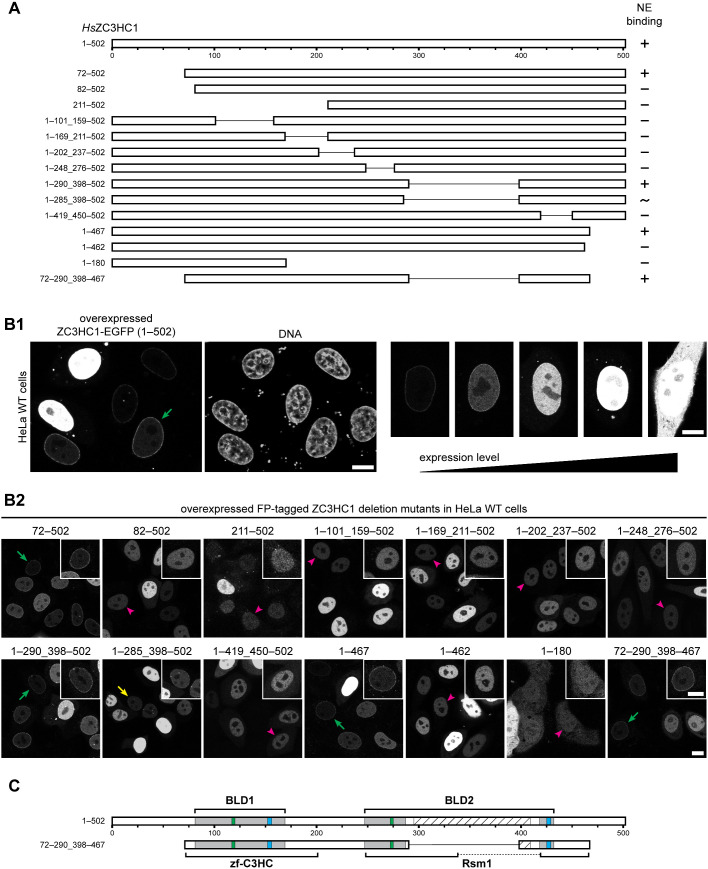
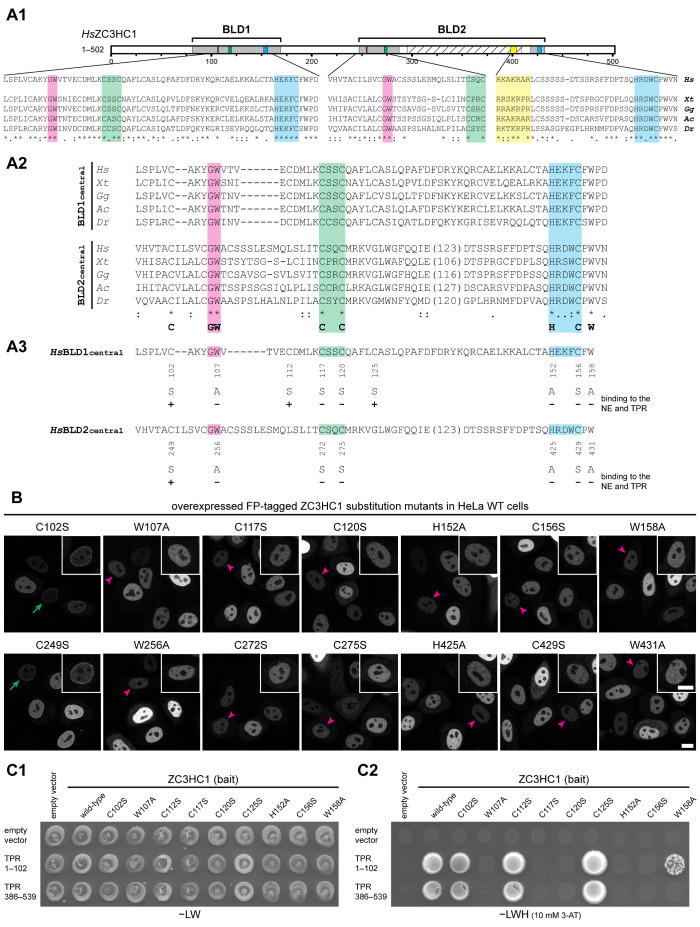
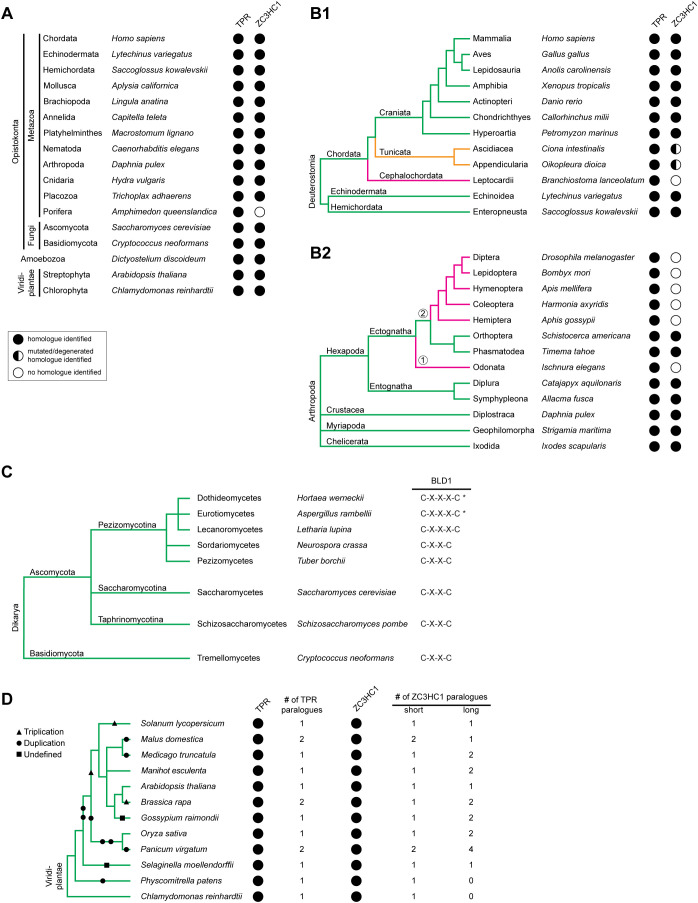
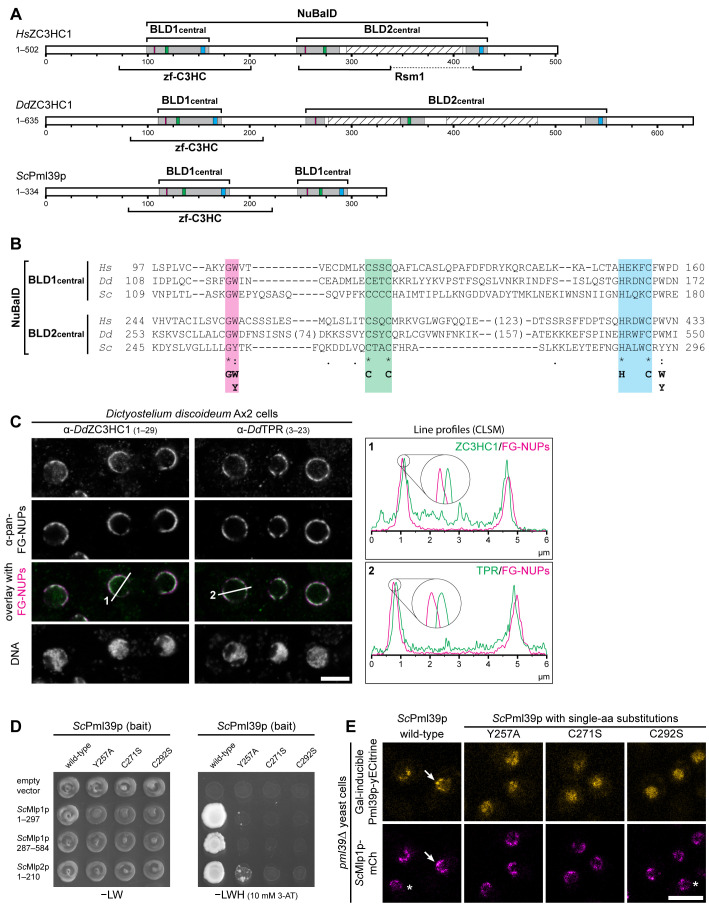
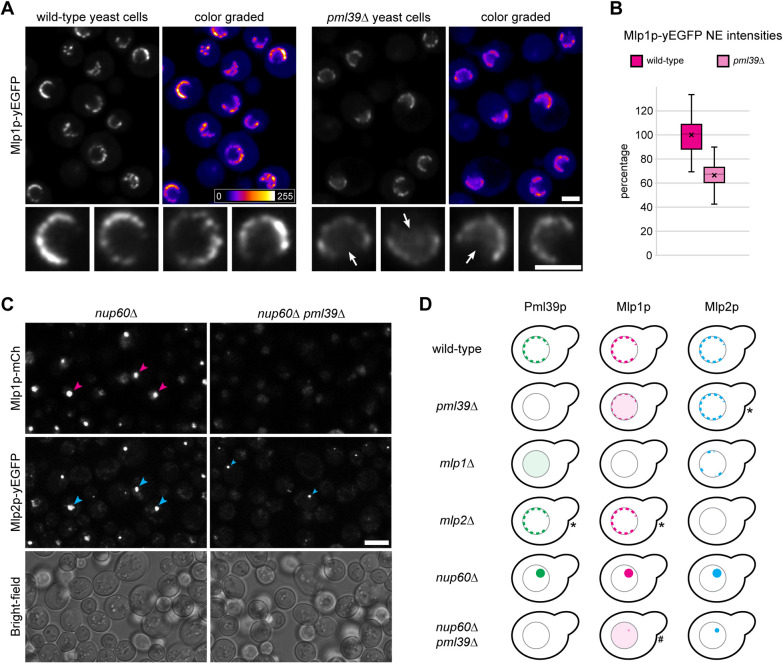
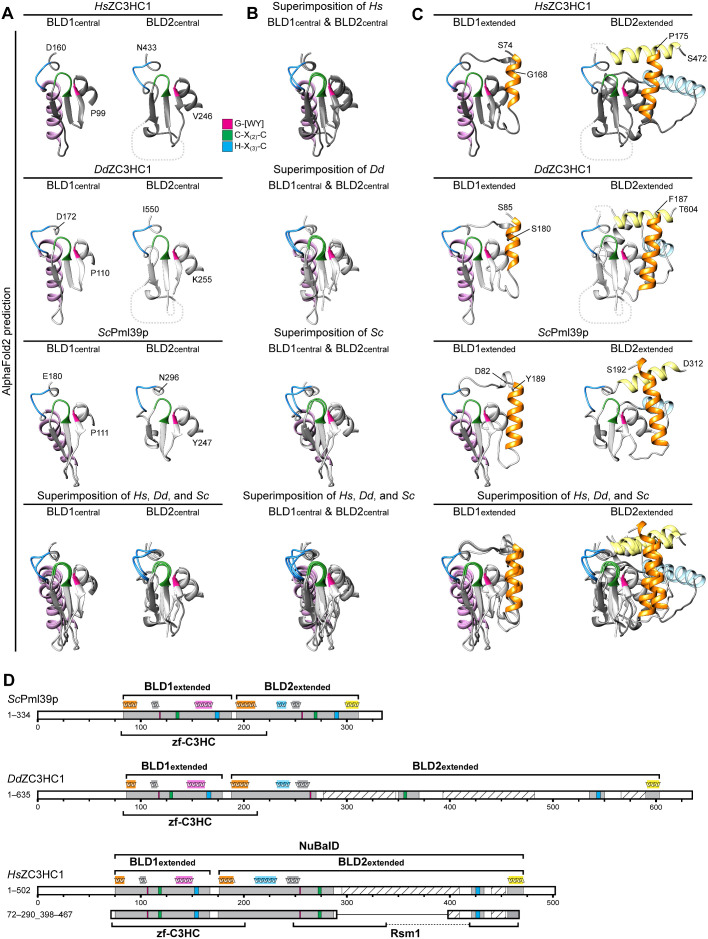
The proteins ZC3HC1 and TPR are structural components of the nuclear basket (NB), a fibrillar structure attached to the nucleoplasmic side of the nuclear pore complex (NPC). ZC3HC1 initially binds to the NB in a TPR-dependent manner and can subsequently recruit additional TPR polypeptides to this structure. Here, we examined the molecular properties of ZC3HC1 that enable its initial binding to the NB and TPR. We report the identification and definition of a nuclear basket-interaction domain (NuBaID) of HsZC3HC1 that comprises two similarly built modules, both essential for binding the NB-resident TPR. We show that such a bimodular construction is evolutionarily conserved, which we further investigated in Dictyostelium discoideum and Saccharomyces cerevisiae. Presenting ScPml39p as the ZC3HC1 homologue in budding yeast, we show that the bimodular NuBaID of Pml39p is essential for binding to the yeast NB and its TPR homologues ScMlp1p and ScMlp2p, and we further demonstrate that Pml39p enables linkage between subpopulations of Mlp1p. We eventually delineate the common NuBaID of the human, amoebic, and yeast homologue as the defining structural entity of a unique protein not found in all but likely present in most taxa of the eukaryotic realm.
Figures
References
-
- Aherrahrou R, Reinberger T, Werner J, Otto M, Al-Hasani J, Munoz-Venegas ML, Civelek M, Schunkert H, Kessler T, Erdmann J, et al. (2021). Deficiency of ZC3HC1 increases vascular smooth muscle cell migration, proliferation and neointima formation following injury. bioRxiv 10.1101/2021.09.29.462212. - DOI
-
- Ashkenazy-Titelman A, Shav-Tal Y, Kehlenbach RH (2020). Into the basket and beyond: the journey of mRNA through the nuclear pore complex. Biochem J 477, 23–44. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
