

Limiting Mrs2-dependent mitochondrial Mg2+ uptake induces metabolic programming in prolonged dietary stress
- PMID: 36857182
- PMCID: PMC10134742
- DOI: 10.1016/j.celrep.2023.112155
Limiting Mrs2-dependent mitochondrial Mg2+ uptake induces metabolic programming in prolonged dietary stress
Abstract
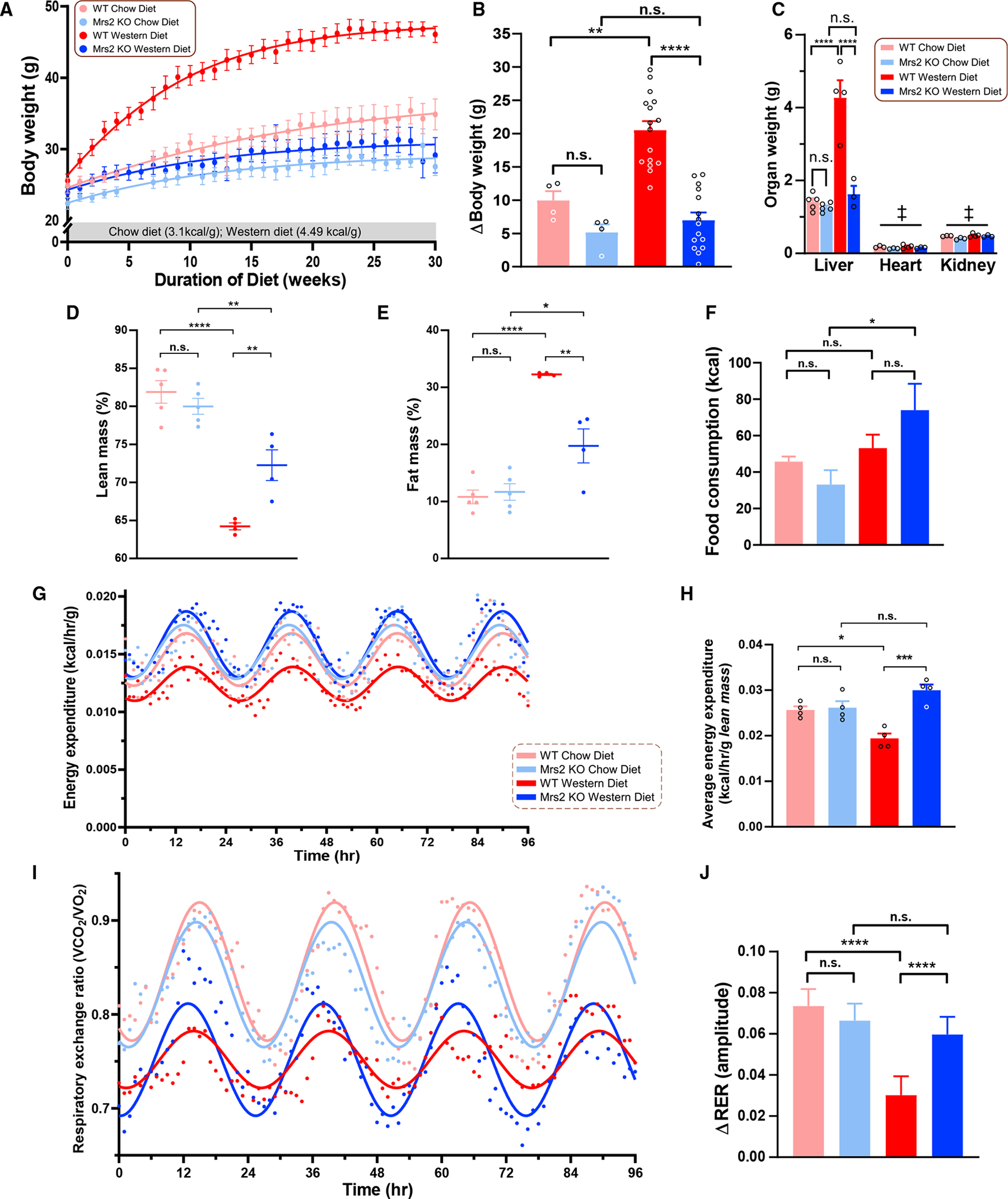
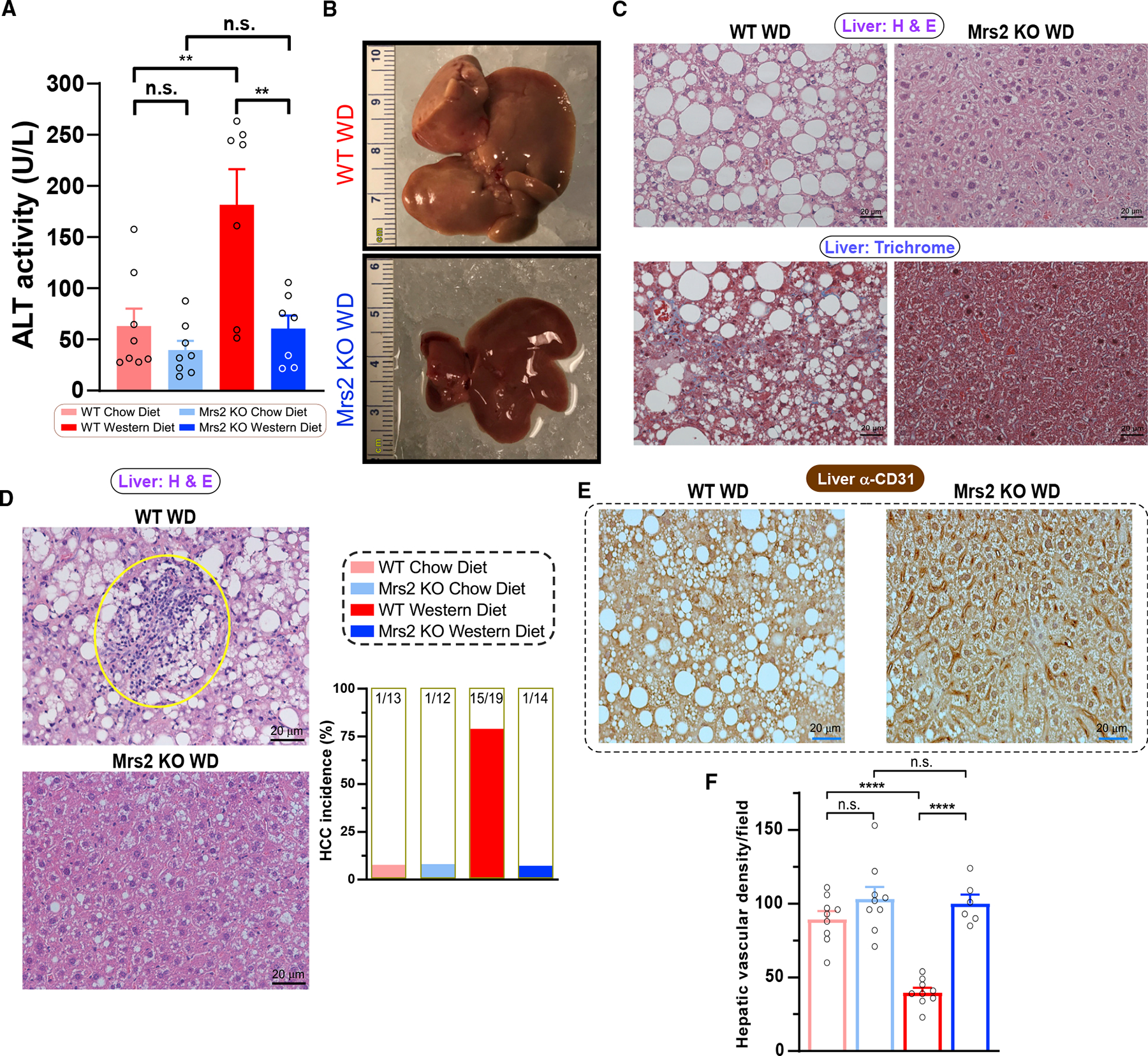
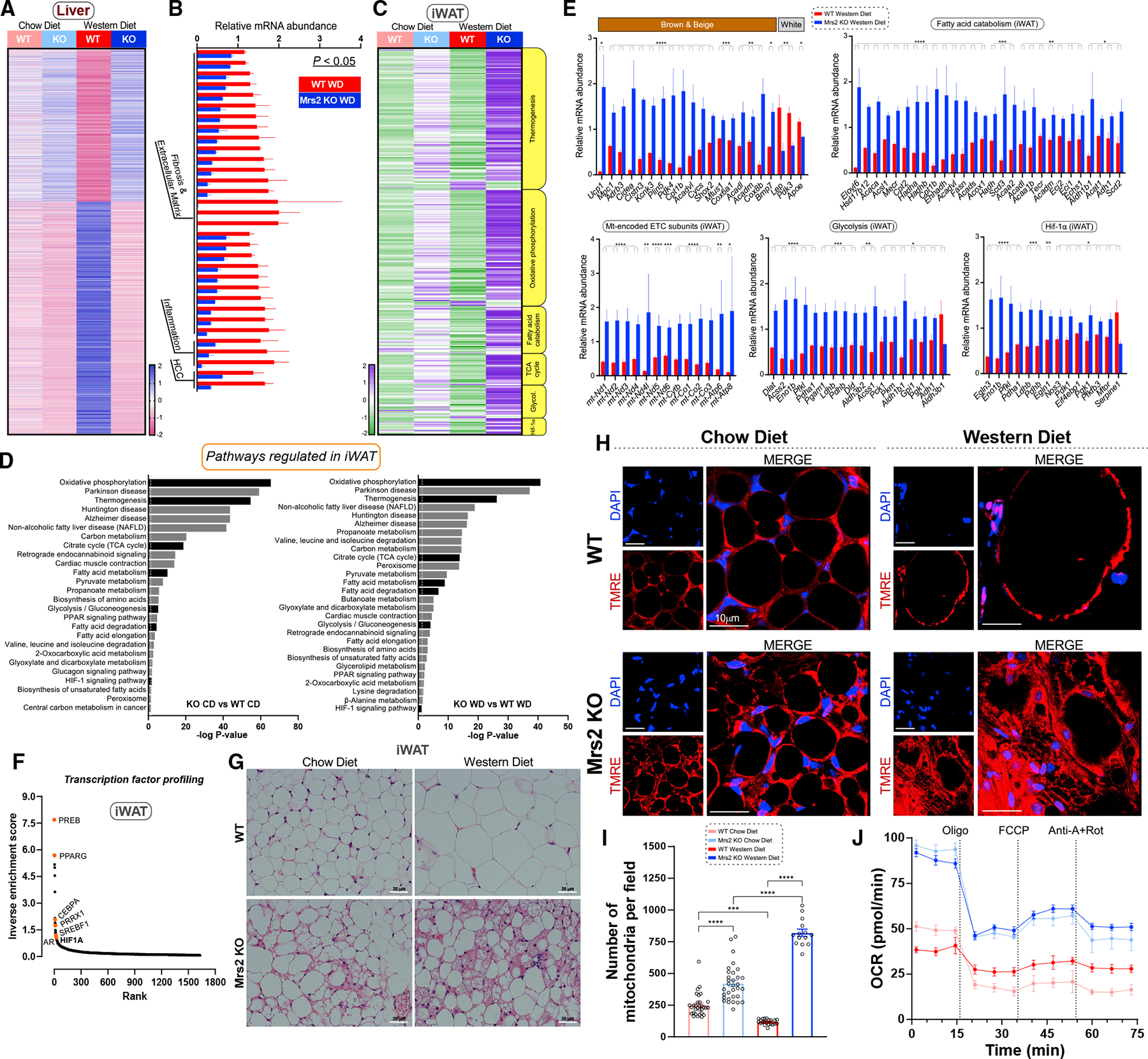
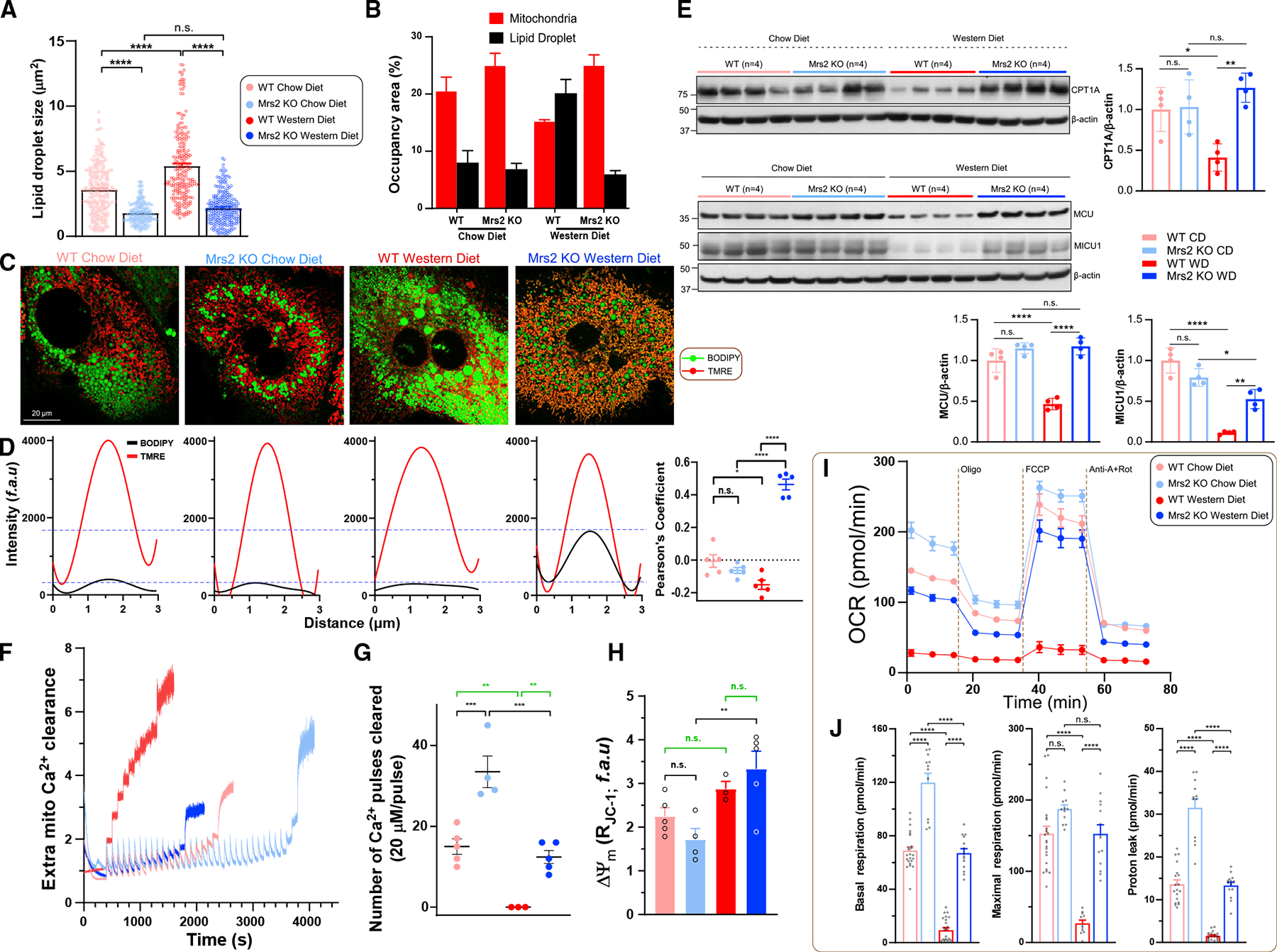
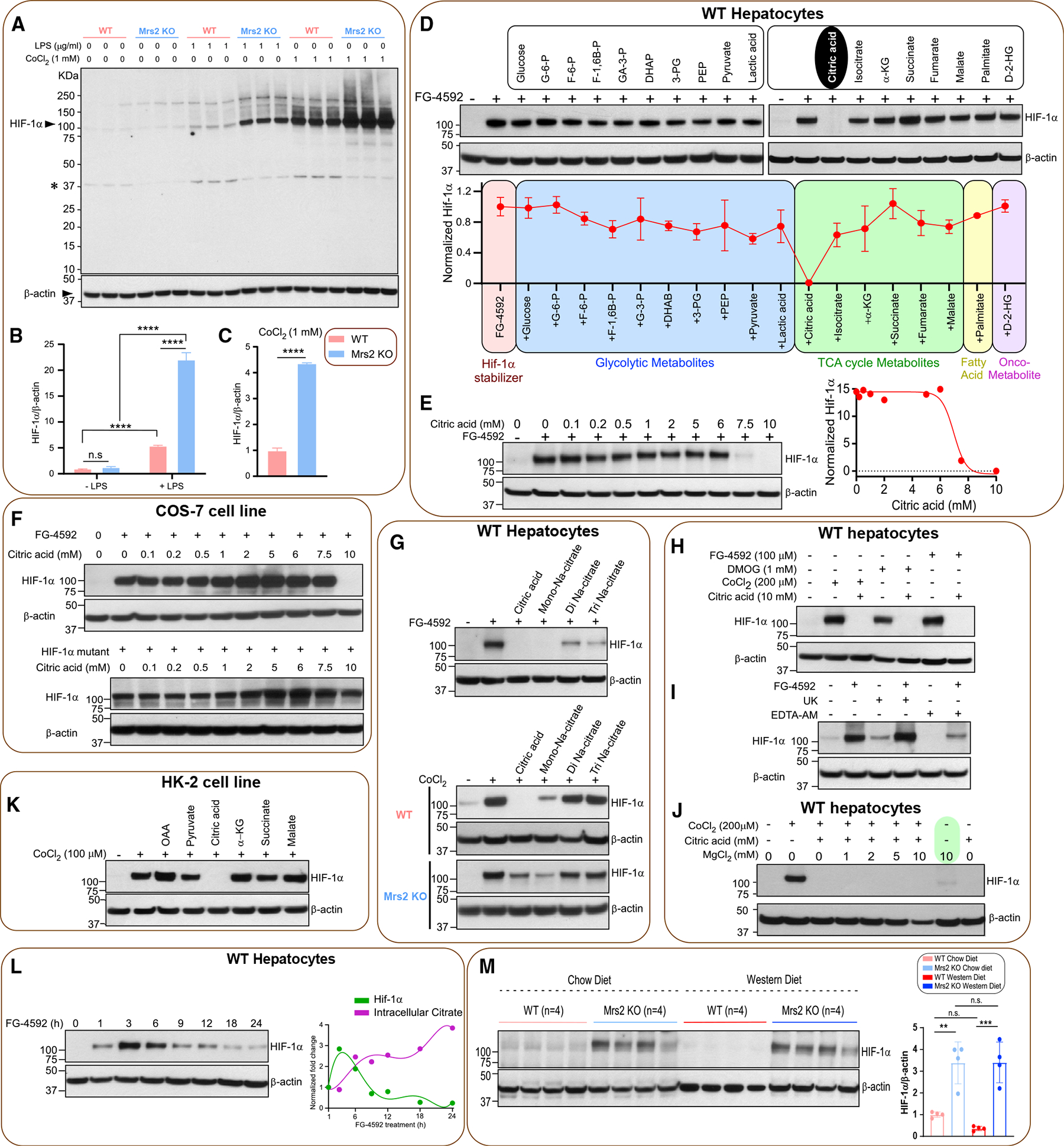
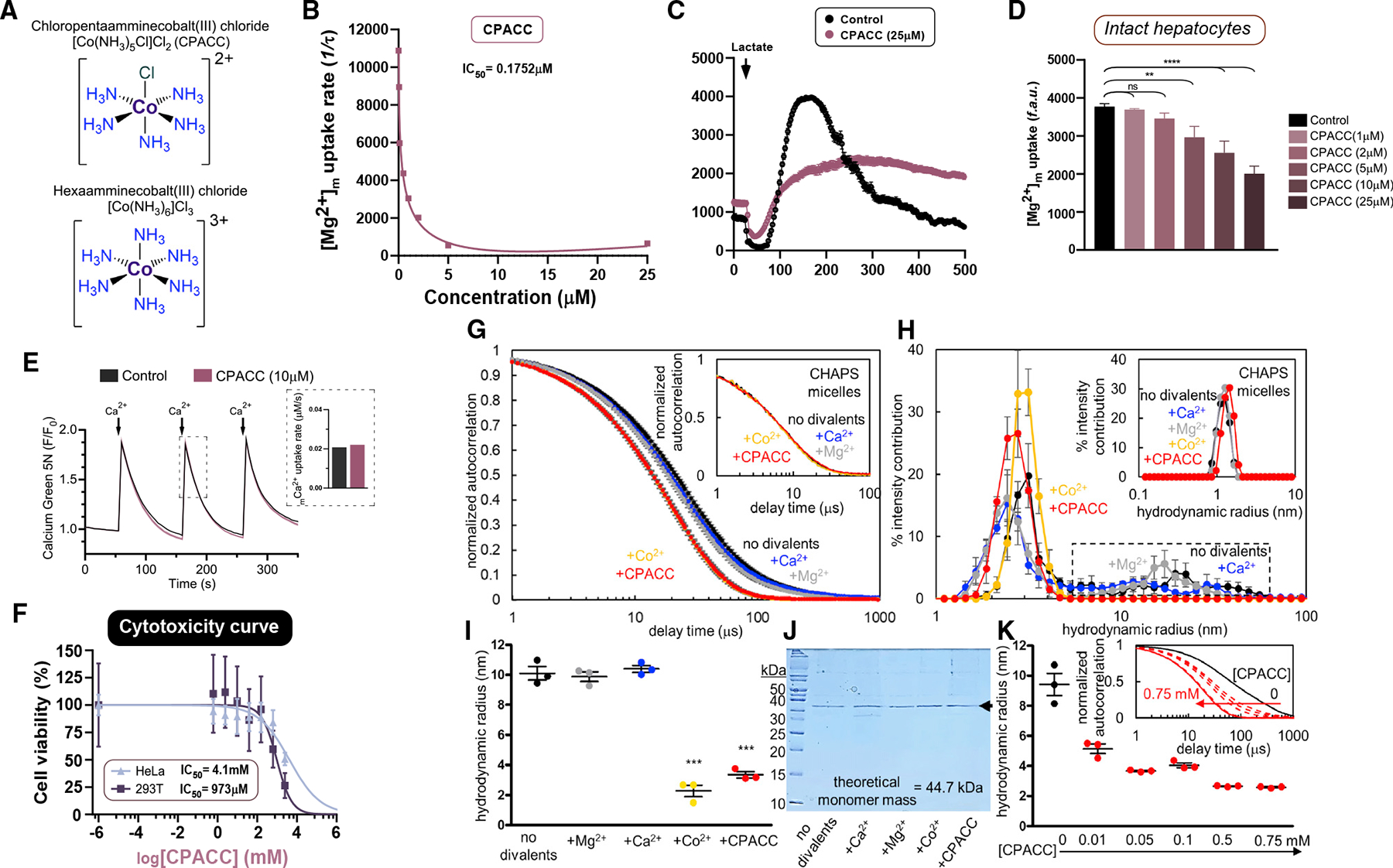
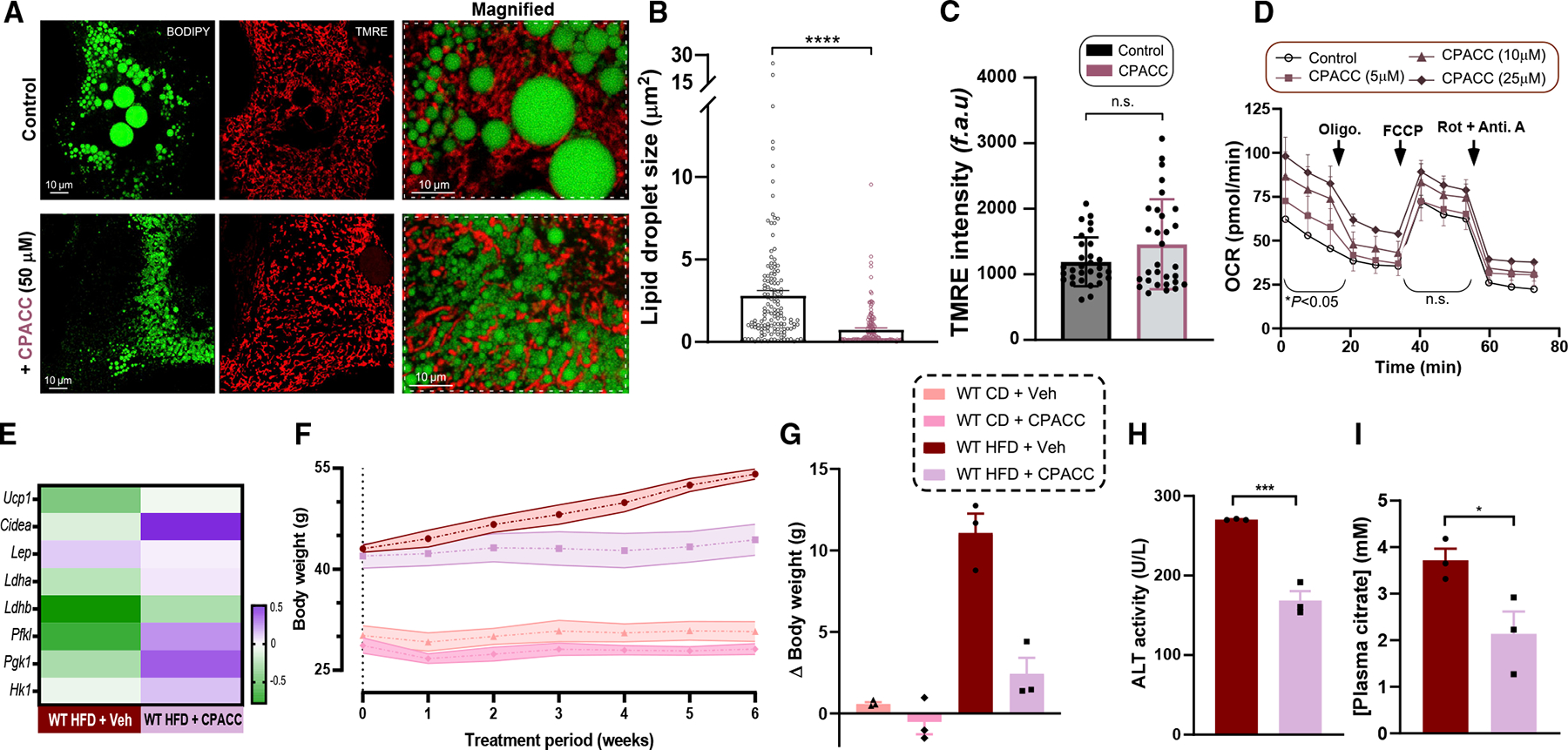
The most abundant cellular divalent cations, Mg2+ (mM) and Ca2+ (nM-μM), antagonistically regulate divergent metabolic pathways with several orders of magnitude affinity preference, but the physiological significance of this competition remains elusive. In mice consuming a Western diet, genetic ablation of the mitochondrial Mg2+ channel Mrs2 prevents weight gain, enhances mitochondrial activity, decreases fat accumulation in the liver, and causes prominent browning of white adipose. Mrs2 deficiency restrains citrate efflux from the mitochondria, making it unavailable to support de novo lipogenesis. As citrate is an endogenous Mg2+ chelator, this may represent an adaptive response to a perceived deficit of the cation. Transcriptional profiling of liver and white adipose reveals higher expression of genes involved in glycolysis, β-oxidation, thermogenesis, and HIF-1α-targets, in Mrs2-/- mice that are further enhanced under Western-diet-associated metabolic stress. Thus, lowering mMg2+ promotes metabolism and dampens diet-induced obesity and metabolic syndrome.
Keywords: CP: Metabolism; HCC; HIF1; MCU; Mrs2; NAFLD; Western diet; adipose expansion; adipose tissue; calcium channel; cardiometabolic disease; diabetes; energy imbalance; hepatocytes; liver; magnesium channel; metabolic disease; metabolic syndrome; mitochondrial dysfunction; obesity; whole-body metabolism.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.M. is an inventor on a patent filed by UTHSA on CPACC as a Mrs2 blocker for Mg(2+) in physiology and disease. J.A.B. is a consultant to Pfizer.
Figures
Comment in
-
Decreasing magnesium reverses the effects of Western Diet.Lab Anim (NY). 2023 May;52(5):101. doi: 10.1038/s41684-023-01172-7. Lab Anim (NY). 2023. PMID: 37137986 No abstract available.
References
-
- Tomar D, Jaña F, Dong Z, Quinn WJ 3rd, Jadiya P, Breves SL, Daw CC, Srikantan S, Shanmughapriya S, Nemani N, et al. (2019). Blockade of MCU-mediated Ca(2+) uptake perturbs lipid metabolism via PP4-dependent AMPK dephosphorylation. Cell Rep. 26, 3709–3725.e7. 10.1016/j.celrep.2019.02.107. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
