

The Effects of Sodium Propionate Supplementation in the Diet with High Soybean Meal on Growth Performance, Intestinal Health, and Immune Resistance to Bacterial Infection in Turbot (Scophthalmus maximus L.)
- PMID: 36860468
- PMCID: PMC9973224
- DOI: 10.1155/2022/8952755
The Effects of Sodium Propionate Supplementation in the Diet with High Soybean Meal on Growth Performance, Intestinal Health, and Immune Resistance to Bacterial Infection in Turbot (Scophthalmus maximus L.)
Abstract
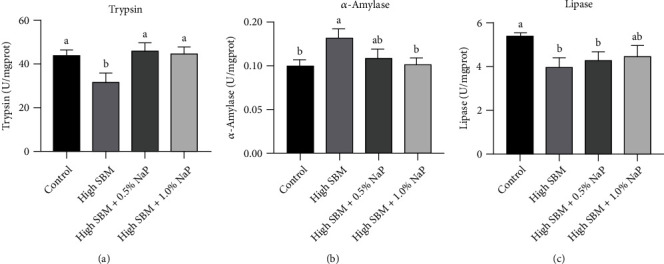
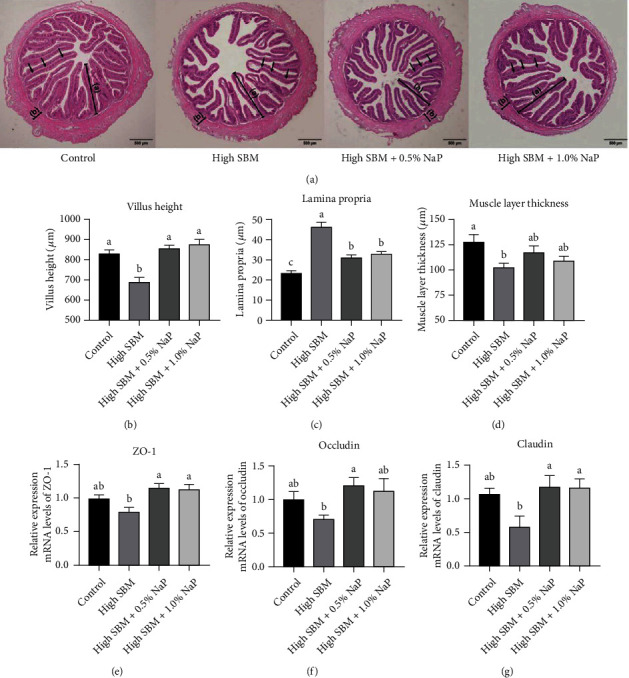
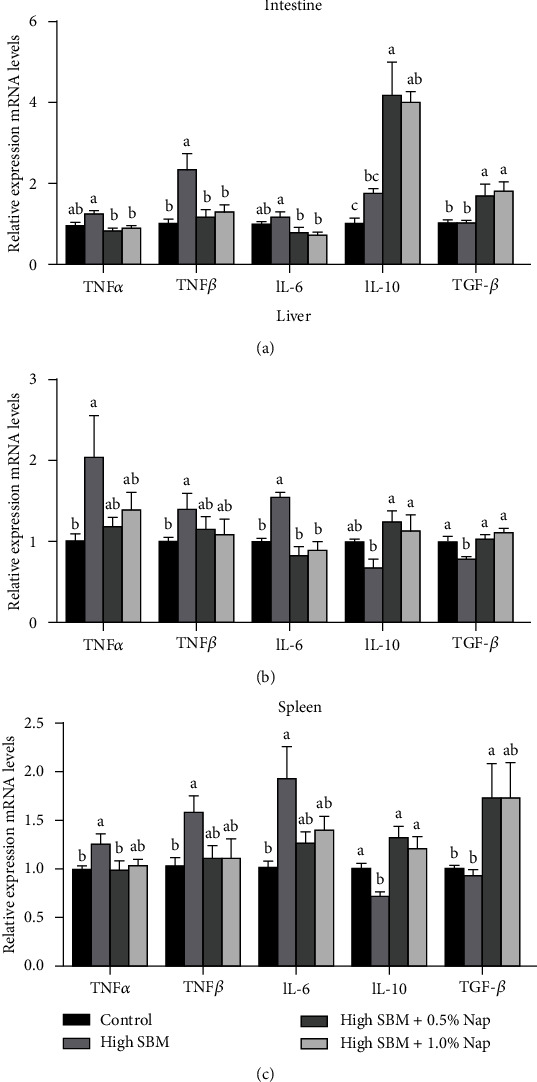
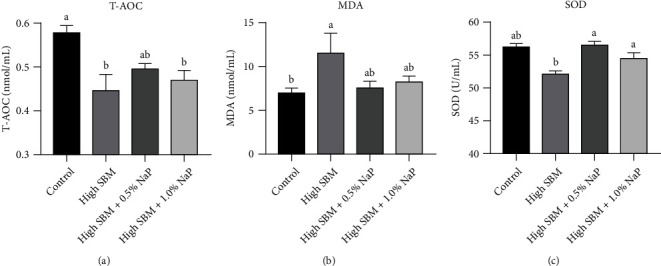
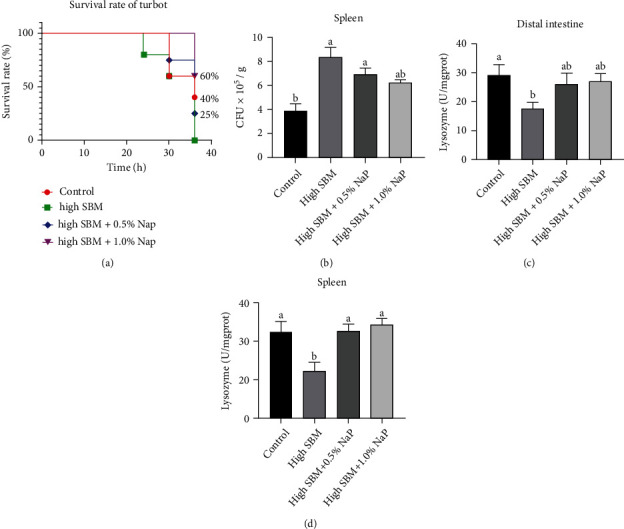
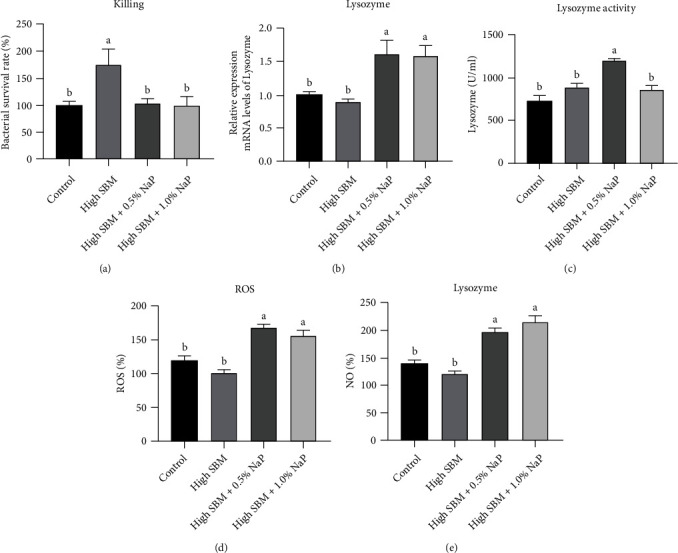
Short-chain fatty acids (SCFAs) are the products of the microbial fermentation of dietary fiber in the intestine. Acetate, propionate, and butyrate are the most abundant SCFA metabolites and play an important role in maintaining host health. This study was aimed at investigating the effects of sodium propionate (NaP) supplementation in the diet with a high proportion of soybean meal (SBM) on the growth, inflammatory status, and anti-infectious ability in juvenile turbot. Four experimental diets were designed: (1) fish meal- (FM-) based diet (control group), (2) SBM protein replacing 45% FM protein in the diet (high SBM group), (3) 0.5% NaP supplementation in the high SBM diet (high SBM+0.5% NaP group), and (4) 1.0% NaP supplementation in the high SBM diet (high SBM+1.0% NaP group). The results confirmed that the fish fed the high SBM diet for 8 weeks showed the decreased growth performance, the typical enteritis symptoms, and the increased mortality responding to Edwardsiella tarda (E. tarda) infection. However, 0.5% NaP supplementation in the high SBM diet promoted the growth performance of turbot and restored the activities of digestive enzymes in the intestine. Moreover, dietary NaP ameliorated the intestinal morphology, enhanced the expression of intestinal tight junction proteins, improved the antioxidant capacity, and suppressed the inflammatory status in turbot. Finally, the expression of antibacterial components and the resistance to bacterial infection were increased in NaP-fed turbot, especially in high SBM+1.0% NaP group. In conclusion, the supplementation of NaP in high SBM diet promotes the growth and health in turbot and provides a theoretical basis for the development of NaP as a functional additive in fish feed.
Copyright © 2022 Huiyuan Sun et al.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
The role of sodium butyrate in modulating growth, intestinal health, and antimicrobial efficacy in turbot (Scophthalmus maximus L.) fed high soy diets.Sci Rep. 2024 Dec 30;14(1):32033. doi: 10.1038/s41598-024-83704-w. Sci Rep. 2024. PMID: 39739006 Free PMC article.
-
Sodium butyrate supplementation in high-soybean meal diets for turbot (Scophthalmus maximus L.): Effects on inflammatory status, mucosal barriers and microbiota in the intestine.Fish Shellfish Immunol. 2019 May;88:65-75. doi: 10.1016/j.fsi.2019.02.064. Epub 2019 Mar 3. Fish Shellfish Immunol. 2019. PMID: 30840856
-
Effects of dietary raw or Enterococcus faecium fermented soybean meal on growth, antioxidant status, intestinal microbiota, morphology, and inflammatory responses in turbot (Scophthalmus maximus L.).Fish Shellfish Immunol. 2020 May;100:261-271. doi: 10.1016/j.fsi.2020.02.070. Epub 2020 Mar 3. Fish Shellfish Immunol. 2020. PMID: 32135340
-
Dietary lipid levels affected antioxidative status, inflammation response, apoptosis and microbial community in the intestine of juvenile turbot (Scophthalmus maximus L.).Comp Biochem Physiol A Mol Integr Physiol. 2022 Feb;264:111118. doi: 10.1016/j.cbpa.2021.111118. Epub 2021 Nov 16. Comp Biochem Physiol A Mol Integr Physiol. 2022. PMID: 34793954 Review.
-
Effects of bioprocessed soybean meal and nucleotide supplementation on growth, physiology and histomorphology in largemouth bass, Micropterus salmoides, juveniles.Comp Biochem Physiol A Mol Integr Physiol. 2021 Oct;260:111038. doi: 10.1016/j.cbpa.2021.111038. Epub 2021 Jul 13. Comp Biochem Physiol A Mol Integr Physiol. 2021. PMID: 34271205 Review.
Cited by
-
Exploring Antibacterial Activity of Fish Protein Hydrolysate In Vitro Against Vibrio Strains and Disease Resistance to V. harveyi in Turbot (Scophthalmus maximus).Aquac Nutr. 2025 Jan 28;2025:3446155. doi: 10.1155/anu/3446155. eCollection 2025. Aquac Nutr. 2025. PMID: 39949355 Free PMC article.
-
Dietary High Glycinin Reduces Growth Performance and Impairs Liver and Intestinal Health Status of Orange-Spotted Grouper (Epinephelus coioides).Animals (Basel). 2023 Aug 12;13(16):2605. doi: 10.3390/ani13162605. Animals (Basel). 2023. PMID: 37627396 Free PMC article.
-
Probiotic Pediococcus pentosaceus restored gossypol-induced intestinal barrier injury by increasing propionate content in Nile tilapia.J Anim Sci Biotechnol. 2024 Apr 7;15(1):54. doi: 10.1186/s40104-024-01011-w. J Anim Sci Biotechnol. 2024. PMID: 38582865 Free PMC article.
References
-
- El-Adawy M., El-Aziz M. A., El-Shazly K., Ali N. G., El-Magd M. A. Dietary propionic acid enhances antibacterial and immunomodulatory effects of oxytetracycline on Nile tilapia, Oreochromis niloticus. Environmental Science and Pollution Research . 2018;25(34):34200–34211. doi: 10.1007/s11356-018-3206-5. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Research Materials