

A parathyroid hormone/salt-inducible kinase signaling axis controls renal vitamin D activation and organismal calcium homeostasis
- PMID: 36862513
- PMCID: PMC10145948
- DOI: 10.1172/JCI163627
A parathyroid hormone/salt-inducible kinase signaling axis controls renal vitamin D activation and organismal calcium homeostasis
Abstract
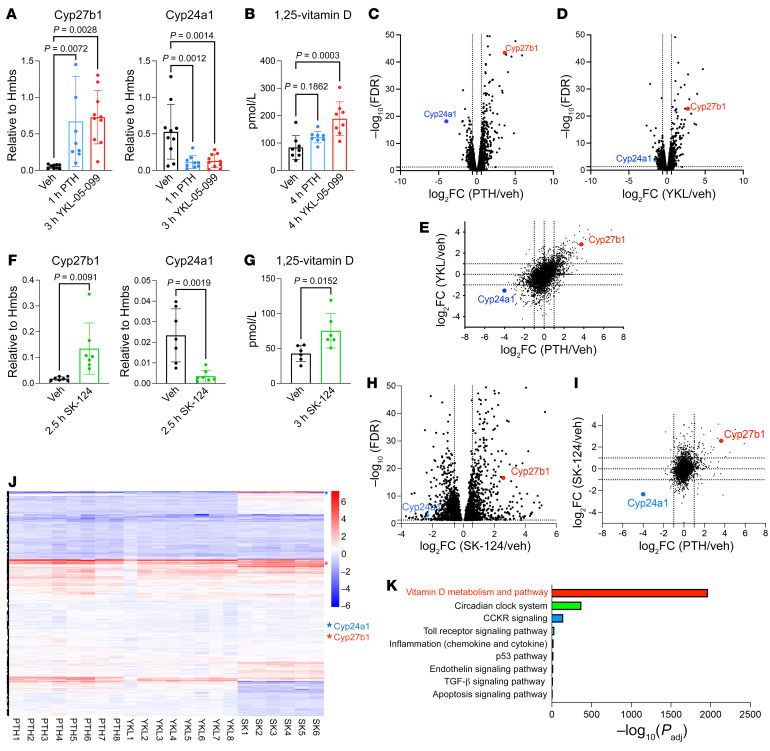
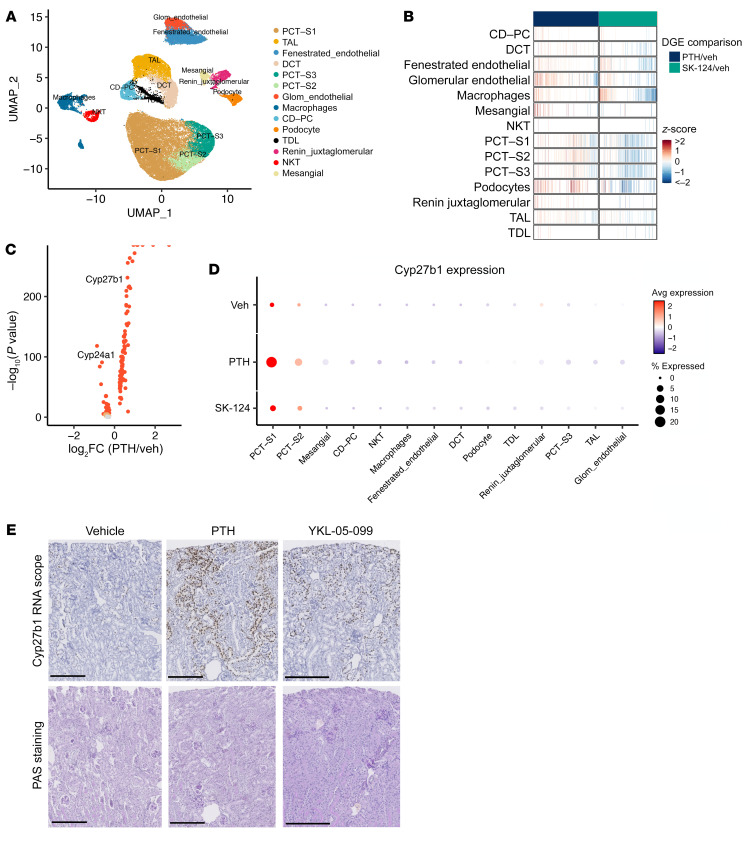
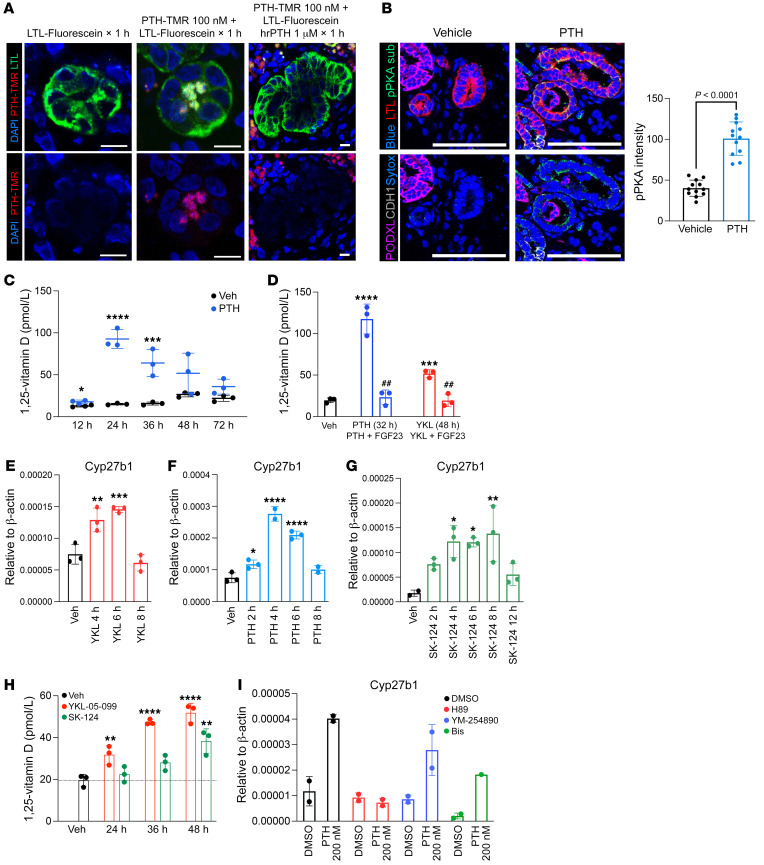
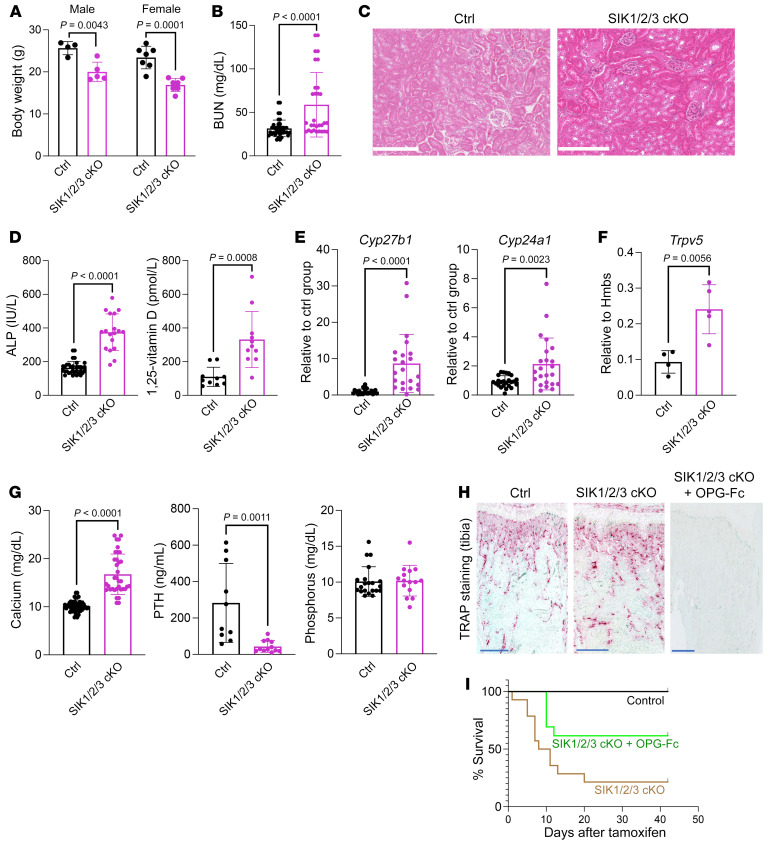
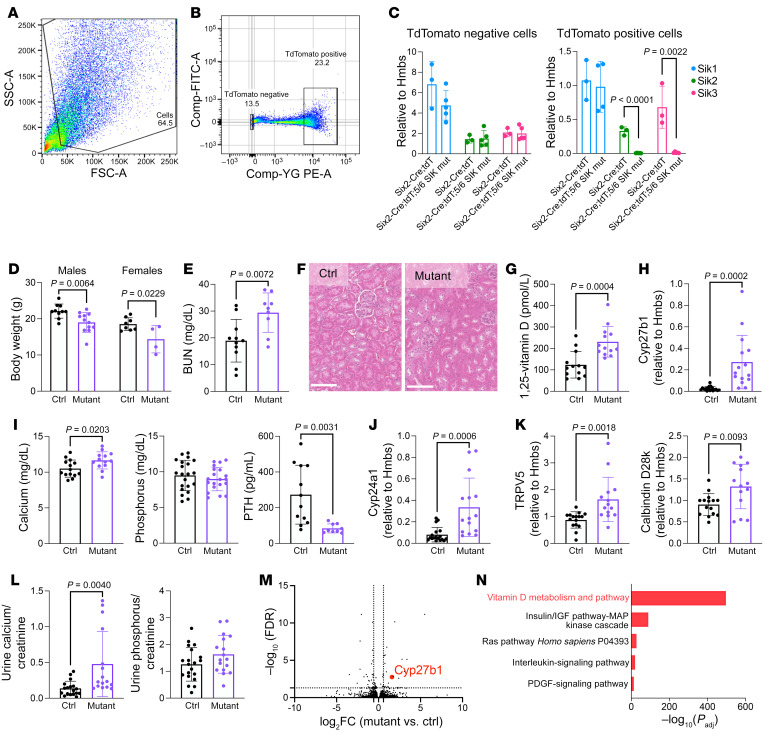
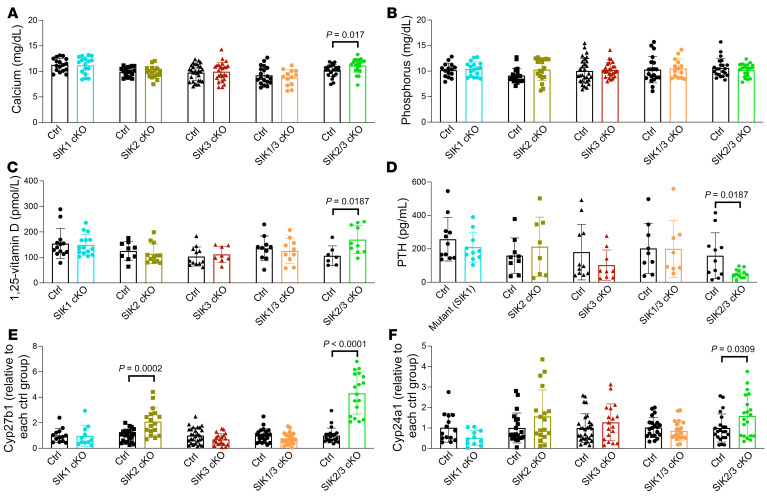
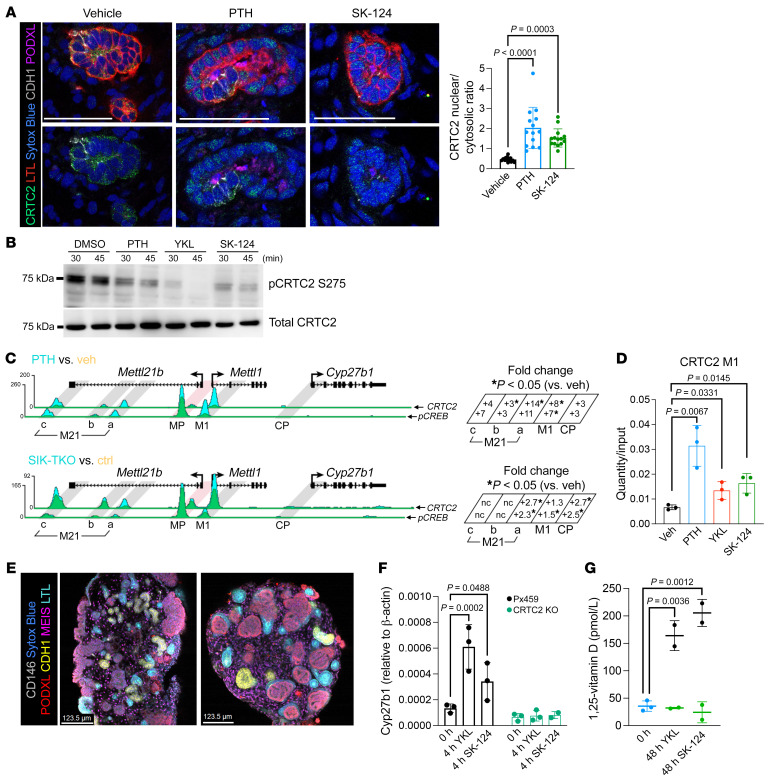
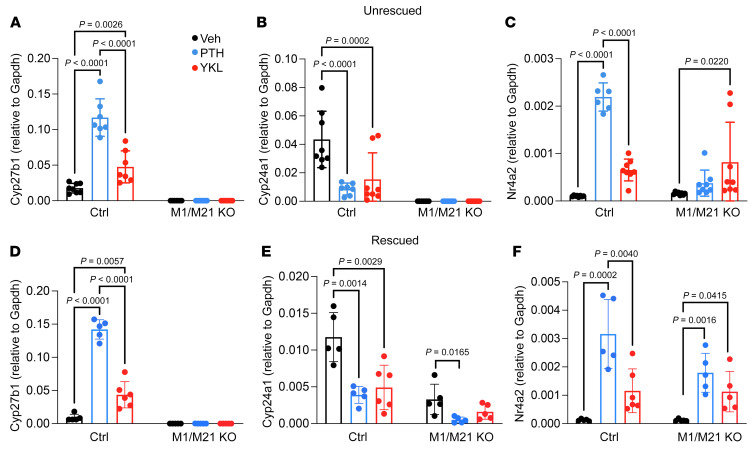
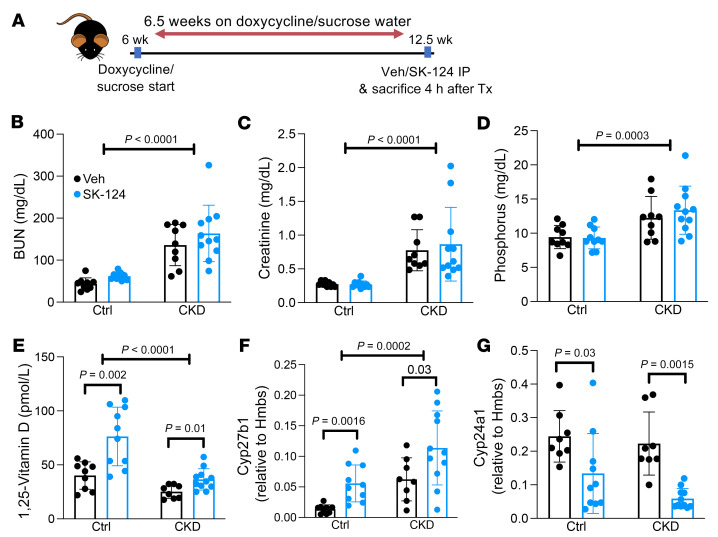
The renal actions of parathyroid hormone (PTH) promote 1,25-vitamin D generation; however, the signaling mechanisms that control PTH-dependent vitamin D activation remain unknown. Here, we demonstrated that salt-inducible kinases (SIKs) orchestrated renal 1,25-vitamin D production downstream of PTH signaling. PTH inhibited SIK cellular activity by cAMP-dependent PKA phosphorylation. Whole-tissue and single-cell transcriptomics demonstrated that both PTH and pharmacologic SIK inhibitors regulated a vitamin D gene module in the proximal tubule. SIK inhibitors increased 1,25-vitamin D production and renal Cyp27b1 mRNA expression in mice and in human embryonic stem cell-derived kidney organoids. Global- and kidney-specific Sik2/Sik3 mutant mice showed Cyp27b1 upregulation, elevated serum 1,25-vitamin D, and PTH-independent hypercalcemia. The SIK substrate CRTC2 showed PTH and SIK inhibitor-inducible binding to key Cyp27b1 regulatory enhancers in the kidney, which were also required for SIK inhibitors to increase Cyp27b1 in vivo. Finally, in a podocyte injury model of chronic kidney disease-mineral bone disorder (CKD-MBD), SIK inhibitor treatment stimulated renal Cyp27b1 expression and 1,25-vitamin D production. Together, these results demonstrated a PTH/SIK/CRTC signaling axis in the kidney that controls Cyp27b1 expression and 1,25-vitamin D synthesis. These findings indicate that SIK inhibitors might be helpful for stimulation of 1,25-vitamin D production in CKD-MBD.
Keywords: Bone Biology; Calcium; G protein–coupled receptors; Nephrology; Protein kinases.
Figures
Comment in
-
Renal proximal tubule cells: power and finesse.J Clin Invest. 2023 May 1;133(9):e169607. doi: 10.1172/JCI169607. J Clin Invest. 2023. PMID: 37115697 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
