

Near-physiological in vitro assembly of 50S ribosomes involves parallel pathways
- PMID: 36864669
- PMCID: PMC10085682
- DOI: 10.1093/nar/gkad082
Near-physiological in vitro assembly of 50S ribosomes involves parallel pathways
Abstract
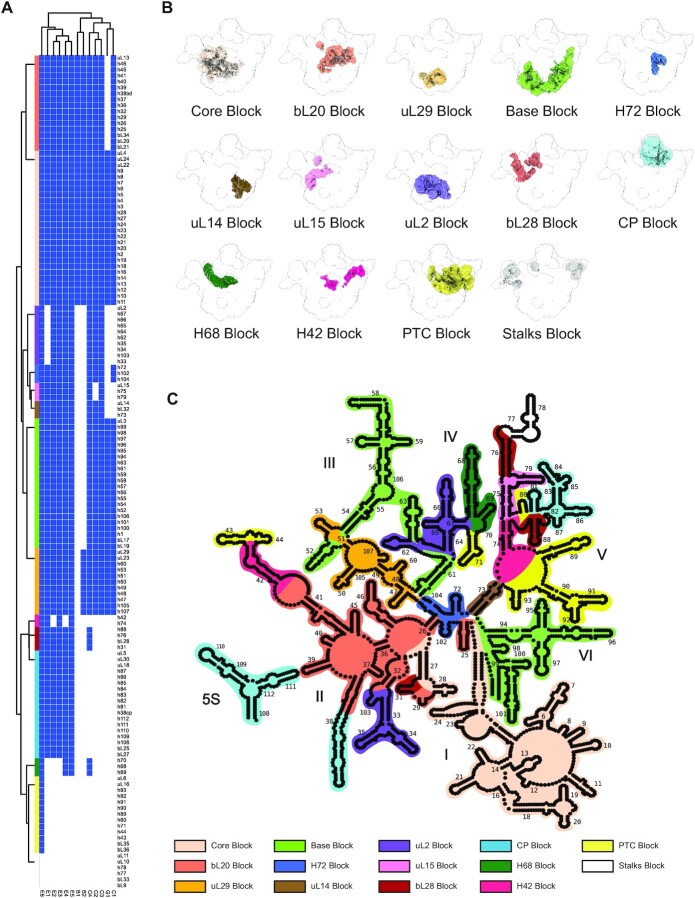
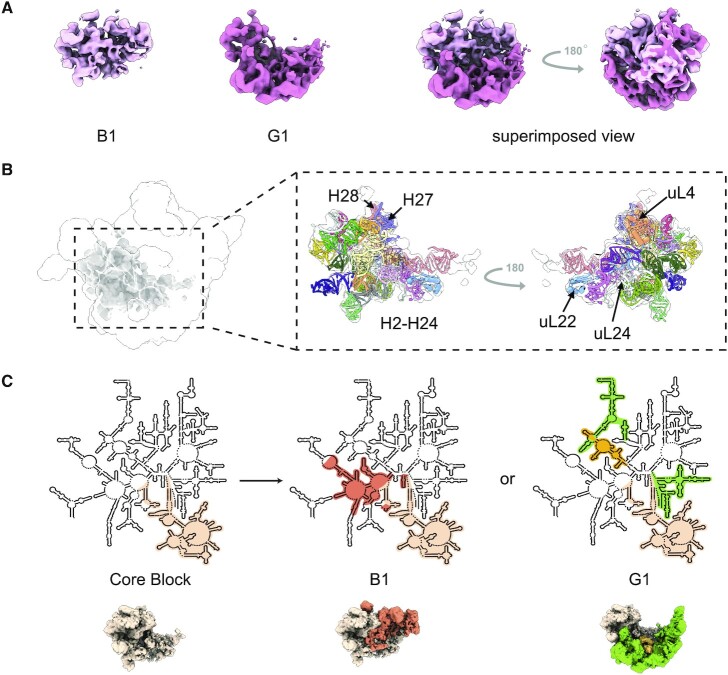
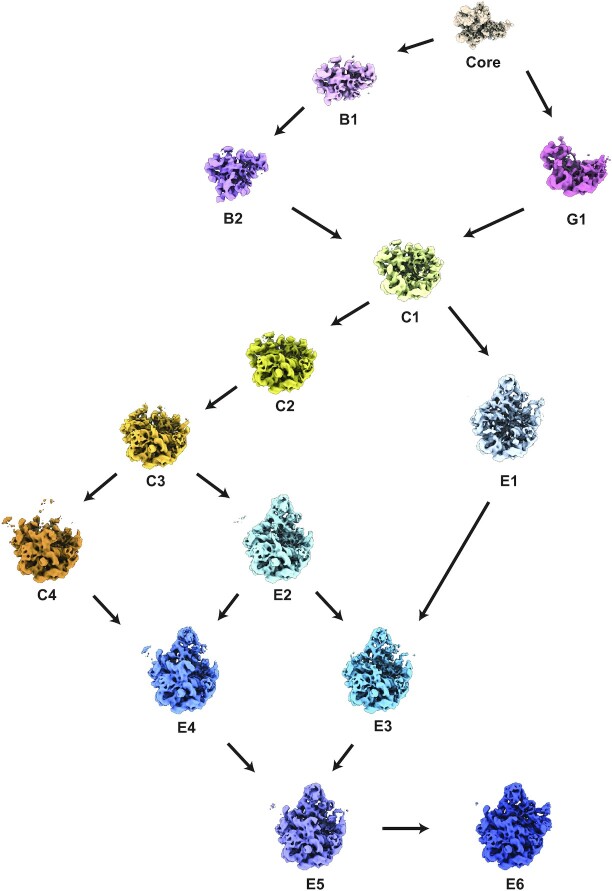
Understanding the assembly principles of biological macromolecular complexes remains a significant challenge, due to the complexity of the systems and the difficulties in developing experimental approaches. As a ribonucleoprotein complex, the ribosome serves as a model system for the profiling of macromolecular complex assembly. In this work, we report an ensemble of large ribosomal subunit intermediate structures that accumulate during synthesis in a near-physiological and co-transcriptional in vitro reconstitution system. Thirteen pre-50S intermediate maps covering the entire assembly process were resolved using cryo-EM single-particle analysis and heterogeneous subclassification. Segmentation of the set of density maps reveals that the 50S ribosome intermediates assemble based on fourteen cooperative assembly blocks, including the smallest assembly core reported to date, which is composed of a 600-nucleotide-long folded rRNA and three ribosomal proteins. The cooperative blocks assemble onto the assembly core following defined dependencies, revealing the parallel pathways at both early and late assembly stages of the 50S subunit.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures




Similar articles
-
Modular Assembly of the Bacterial Large Ribosomal Subunit.Cell. 2016 Dec 1;167(6):1610-1622.e15. doi: 10.1016/j.cell.2016.11.020. Cell. 2016. PMID: 27912064 Free PMC article.
-
Lack of complete cooperativity of ribosome assembly in vitro and its possible relevance to in vivo ribosome assembly and the regulation of ribosomal gene expression.Biochimie. 1991 Jun;73(6):757-67. doi: 10.1016/0300-9084(91)90055-6. Biochimie. 1991. PMID: 1764521
-
Cryo-EM captures early ribosome assembly in action.Nat Commun. 2023 Feb 17;14(1):898. doi: 10.1038/s41467-023-36607-9. Nat Commun. 2023. PMID: 36797249 Free PMC article.
-
Critical steps in the assembly process of the bacterial 50S ribosomal subunit.Nucleic Acids Res. 2024 May 8;52(8):4111-4123. doi: 10.1093/nar/gkae199. Nucleic Acids Res. 2024. PMID: 38554105 Free PMC article. Review.
-
Assembly of bacterial ribosomes.Annu Rev Biochem. 2011;80:501-26. doi: 10.1146/annurev-biochem-062608-160432. Annu Rev Biochem. 2011. PMID: 21529161 Review.
Cited by
-
Autonomous ribosome biogenesis in vitro.Nat Commun. 2025 Jan 8;16(1):514. doi: 10.1038/s41467-025-55853-7. Nat Commun. 2025. PMID: 39779722 Free PMC article.
-
The proline-rich antimicrobial peptide Api137 disrupts large ribosomal subunit assembly and induces misfolding.Nat Commun. 2025 Jan 10;16(1):567. doi: 10.1038/s41467-025-55836-8. Nat Commun. 2025. PMID: 39794318 Free PMC article.
-
Ribosomal RNA modification enzymes stimulate large ribosome subunit assembly in E. coli.Nucleic Acids Res. 2024 Jun 24;52(11):6614-6628. doi: 10.1093/nar/gkae222. Nucleic Acids Res. 2024. PMID: 38554109 Free PMC article.
-
A roadmap for ribosome assembly in human mitochondria.Nat Struct Mol Biol. 2024 Dec;31(12):1898-1908. doi: 10.1038/s41594-024-01356-w. Epub 2024 Jul 11. Nat Struct Mol Biol. 2024. PMID: 38992089 Free PMC article.
-
Putting It All Together: The Roles of Ribosomal Proteins in Nucleolar Stages of 60S Ribosomal Assembly in the Yeast Saccharomyces cerevisiae.Biomolecules. 2024 Aug 9;14(8):975. doi: 10.3390/biom14080975. Biomolecules. 2024. PMID: 39199362 Free PMC article. Review.
References
-
- Friesen J.D. Escherichia coli and Salmonella typhimurium: cellular and molecular biology. Science. 1988; 240:1678–1681. - PubMed
-
- Shajani Z., Sykes M.T., Williamson J.R.. Assembly of bacterial ribosomes. Annu. Rev. Biochem. 2011; 80:501–526. - PubMed
-
- Wilson D.N., Nierhaus K.H.. The weird and wonderful world of bacterial ribosome regulation. Crit. Rev. Biochem. Mol. Biol. 2007; 42:187–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
