

This is a preprint.
Myomatrix arrays for high-definition muscle recording
- PMID: 36865176
- PMCID: PMC9980060
- DOI: 10.1101/2023.02.21.529200
Myomatrix arrays for high-definition muscle recording
Update in
-
Myomatrix arrays for high-definition muscle recording.Elife. 2023 Dec 19;12:RP88551. doi: 10.7554/eLife.88551. Elife. 2023. PMID: 38113081 Free PMC article.
Abstract
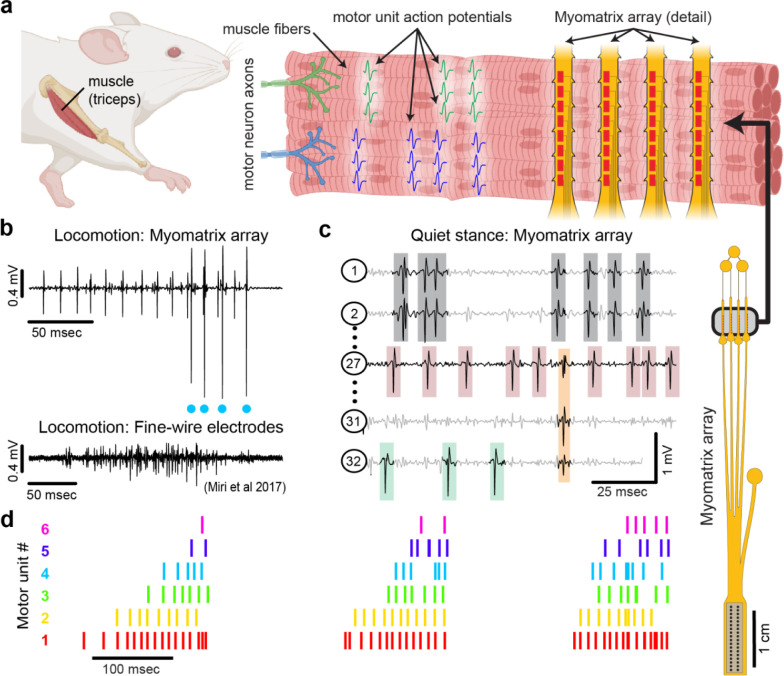
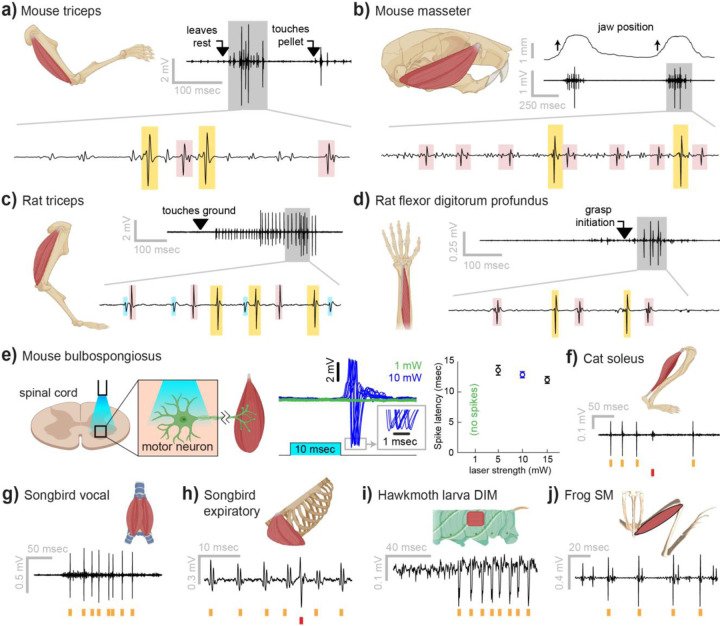
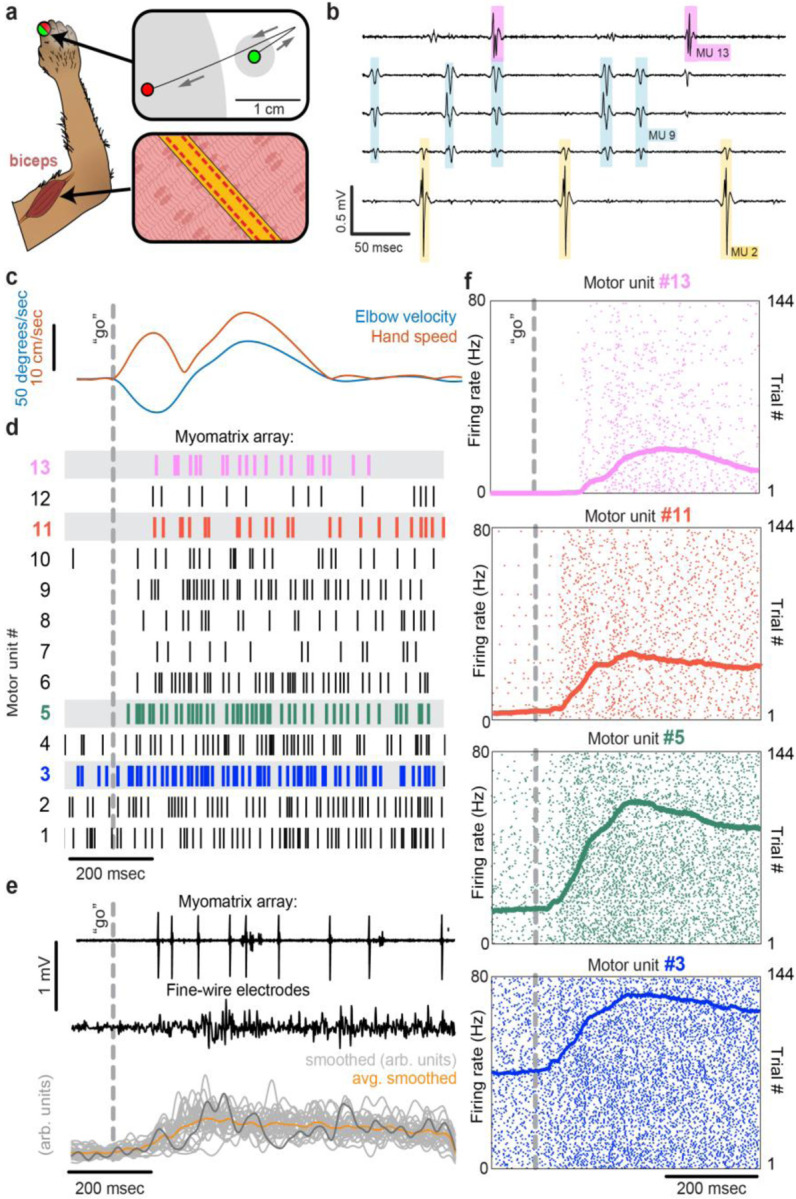
Neurons coordinate their activity to produce an astonishing variety of motor behaviors. Our present understanding of motor control has grown rapidly thanks to new methods for recording and analyzing populations of many individual neurons over time. In contrast, current methods for recording the nervous system's actual motor output - the activation of muscle fibers by motor neurons - typically cannot detect the individual electrical events produced by muscle fibers during natural behaviors and scale poorly across species and muscle groups. Here we present a novel class of electrode devices ("Myomatrix arrays") that record muscle activity at unprecedented resolution across muscles and behaviors. High-density, flexible electrode arrays allow for stable recordings from the muscle fibers activated by a single motor neuron, called a "motor unit", during natural behaviors in many species, including mice, rats, primates, songbirds, frogs, and insects. This technology therefore allows the nervous system's motor output to be monitored in unprecedented detail during complex behaviors across species and muscle morphologies. We anticipate that this technology will allow rapid advances in understanding the neural control of behavior and in identifying pathologies of the motor system.
Figures
References
-
- Pachitariu M., Sridhar S. & Stringer C. Solving the spike sorting problem with Kilosort. bioRxiv, 2023.2001.2007.523036 (2023). 10.1101/2023.01.07.523036 - DOI
-
- Lenschow C. et al. A galanin-positive population of lumbar spinal cord neurons modulates sexual behavior and arousal. bioRxiv, 2022.2010.2004.510783 (2022). 10.1101/2022.10.04.510783 - DOI
Publication types
Associated data
Grants and funding
- U24 NS126936/NS/NINDS NIH HHS/United States
- U19 NS104649/NS/NINDS NIH HHS/United States
- DP2 NS105555/NS/NINDS NIH HHS/United States
- R01 EB022872/EB/NIBIB NIH HHS/United States
- K99 NS126307/NS/NINDS NIH HHS/United States
- R01 NS099375/NS/NINDS NIH HHS/United States
- R01 NS124820/NS/NINDS NIH HHS/United States
- R01 NS104194/NS/NINDS NIH HHS/United States
- R01 NS111479/NS/NINDS NIH HHS/United States
- DP2 NS127291/NS/NINDS NIH HHS/United States
- R35 NS111643/NS/NINDS NIH HHS/United States
- R01 NS084844/NS/NINDS NIH HHS/United States
- RF1 NS128898/NS/NINDS NIH HHS/United States
- R01 NS109237/NS/NINDS NIH HHS/United States
- F32 MH120873/MH/NIMH NIH HHS/United States
- F31 NS124347/NS/NINDS NIH HHS/United States
- U19 NS112959/NS/NINDS NIH HHS/United States
- R01 NS104834/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources