

This is a preprint.
Hypoxia-inducible factor 2 is a key determinant of manganese excess and polycythemia in SLC30A10 deficiency
- PMID: 36865210
- PMCID: PMC9980069
- DOI: 10.1101/2023.02.20.529270
Hypoxia-inducible factor 2 is a key determinant of manganese excess and polycythemia in SLC30A10 deficiency
Update in
-
Hepatic HIF2 is a key determinant of manganese excess and polycythemia in SLC30A10 deficiency.JCI Insight. 2024 Apr 23;9(10):e169738. doi: 10.1172/jci.insight.169738. JCI Insight. 2024. PMID: 38652538 Free PMC article.
Abstract
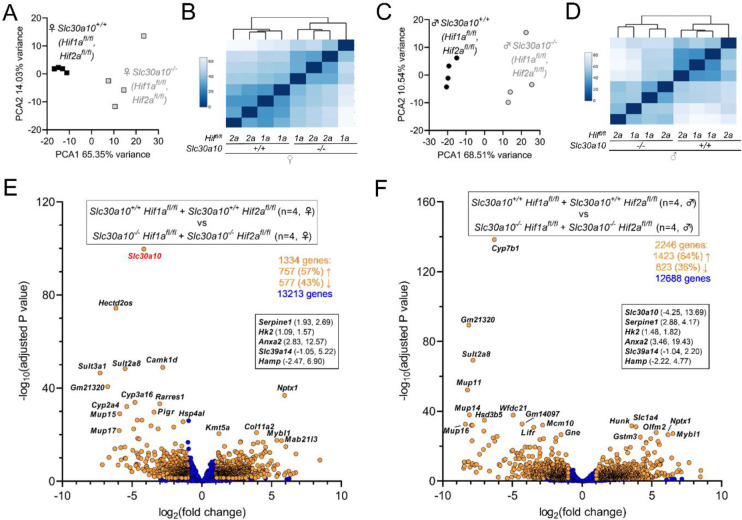
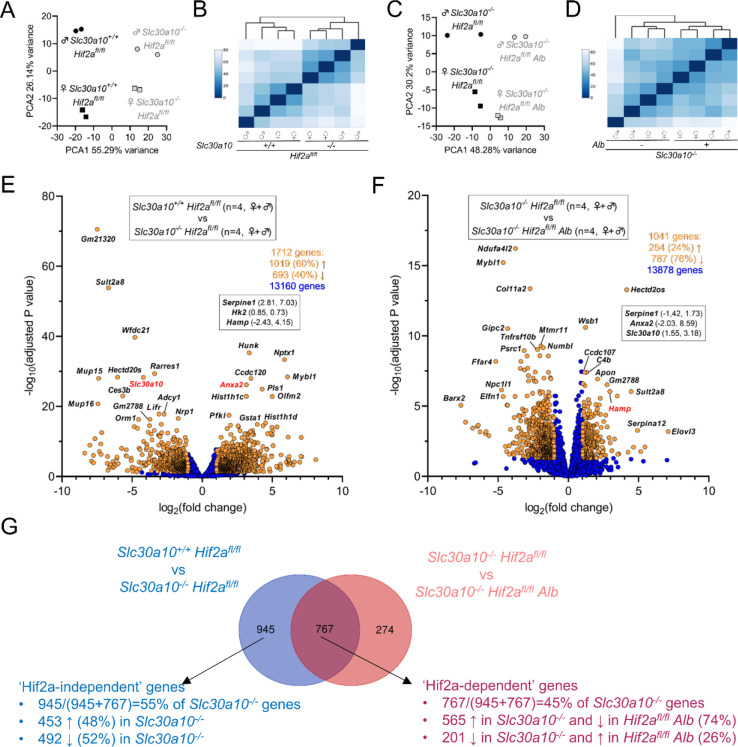
Manganese is an essential yet potentially toxic metal. Initially reported in 2012, mutations in SLC30A10 are the first known inherited cause of manganese excess. SLC30A10 is an apical membrane transport protein that exports manganese from hepatocytes into bile and from enterocytes into the lumen of the gastrointestinal tract. SLC30A10 deficiency results in impaired gastrointestinal manganese excretion, leading to severe manganese excess, neurologic deficits, liver cirrhosis, polycythemia, and erythropoietin excess. Neurologic and liver disease are attributed to manganese toxicity. Polycythemia is attributed to erythropoietin excess, but the basis of erythropoietin excess in SLC30A10 deficiency has yet to be established. Here we demonstrate that erythropoietin expression is increased in liver but decreased in kidneys in Slc30a10-deficient mice. Using pharmacologic and genetic approaches, we show that liver expression of hypoxia-inducible factor 2 (Hif2), a transcription factor that mediates the cellular response to hypoxia, is essential for erythropoietin excess and polycythemia in Slc30a10-deficient mice, while hypoxia-inducible factor 1 (HIF1) plays no discernible role. RNA-seq analysis determined that Slc30a10-deficient livers exhibit aberrant expression of a large number of genes, most of which align with cell cycle and metabolic processes, while hepatic Hif2 deficiency attenuates differential expression of half of these genes in mutant mice. One such gene downregulated in Slc30a10-deficient mice in a Hif2-dependent manner is hepcidin, a hormonal inhibitor of dietary iron absorption. Our analyses indicate that hepcidin downregulation serves to increase iron absorption to meet the demands of erythropoiesis driven by erythropoietin excess. Finally, we also observed that hepatic Hif2 deficiency attenuates tissue manganese excess, although the underlying cause of this observation is not clear at this time. Overall, our results indicate that HIF2 is a key determinant of pathophysiology in SLC30A10 deficiency.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures








Similar articles
-
Hepatic HIF2 is a key determinant of manganese excess and polycythemia in SLC30A10 deficiency.JCI Insight. 2024 Apr 23;9(10):e169738. doi: 10.1172/jci.insight.169738. JCI Insight. 2024. PMID: 38652538 Free PMC article.
-
Manganese transporter Slc30a10 controls physiological manganese excretion and toxicity.J Clin Invest. 2019 Dec 2;129(12):5442-5461. doi: 10.1172/JCI129710. J Clin Invest. 2019. PMID: 31527311 Free PMC article.
-
AAV-mediated hepatic expression of SLC30A10 and the Thr95Ile variant attenuates manganese excess and other phenotypes in Slc30a10-deficient mice.J Biol Chem. 2024 Mar;300(3):105732. doi: 10.1016/j.jbc.2024.105732. Epub 2024 Feb 8. J Biol Chem. 2024. PMID: 38336290 Free PMC article.
-
A case of dystonia with polycythemia and hypermanganesemia caused by SLC30A10 mutation: a treatable inborn error of manganese metabolism.BMC Pediatr. 2019 Jul 9;19(1):229. doi: 10.1186/s12887-019-1611-7. BMC Pediatr. 2019. PMID: 31288771 Free PMC article. Review.
-
Inherited Manganese Disorders and the Brain: What Neurologists Need to Know.Ann Indian Acad Neurol. 2021 Jan-Feb;24(1):15-21. doi: 10.4103/aian.AIAN_789_20. Epub 2021 Feb 5. Ann Indian Acad Neurol. 2021. PMID: 33911374 Free PMC article. Review.
References
-
- Chen P, Bornhorst J, Aschner M. Manganese metabolism in humans. Front. Biosci. Landmark Ed. 2018;23:1655–1679. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources