

This is a preprint.
A cell state specific metabolic vulnerability to GPX4-dependent ferroptosis in glioblastoma
- PMID: 36865302
- PMCID: PMC9980114
- DOI: 10.1101/2023.02.22.529581
A cell state specific metabolic vulnerability to GPX4-dependent ferroptosis in glioblastoma
Update in
-
A cell state-specific metabolic vulnerability to GPX4-dependent ferroptosis in glioblastoma.EMBO J. 2024 Oct;43(20):4492-4521. doi: 10.1038/s44318-024-00176-4. Epub 2024 Aug 27. EMBO J. 2024. PMID: 39192032 Free PMC article.
Abstract
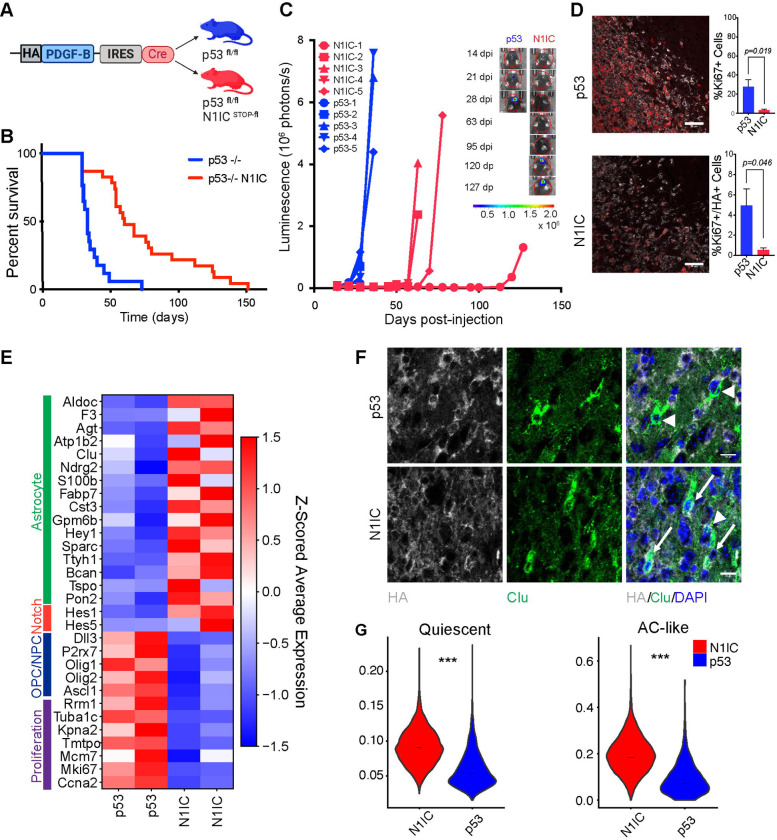
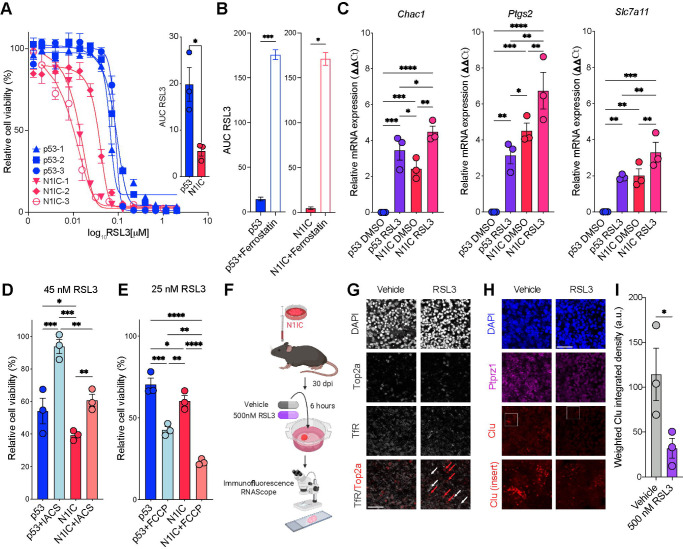
Glioma cells hijack developmental transcriptional programs to control cell state. During neural development, lineage trajectories rely on specialized metabolic pathways. However, the link between tumor cell state and metabolic programs is poorly understood in glioma. Here we uncover a glioma cell state-specific metabolic liability that can be leveraged therapeutically. To model cell state diversity, we generated genetically engineered murine gliomas, induced by deletion of p53 alone (p53) or with constitutively active Notch signaling (N1IC), a pathway critical in controlling cellular fate. N1IC tumors harbored quiescent astrocyte-like transformed cell states while p53 tumors were predominantly comprised of proliferating progenitor-like cell states. N1IC cells exhibit distinct metabolic alterations, with mitochondrial uncoupling and increased ROS production rendering them more sensitive to inhibition of the lipid hydroperoxidase GPX4 and induction of ferroptosis. Importantly, treating patient-derived organotypic slices with a GPX4 inhibitor induced selective depletion of quiescent astrocyte-like glioma cell populations with similar metabolic profiles.
Keywords: Astrocyte; Cell-state; Ferroptosis; Glioblastoma; Metabolism; Quiescent.
Conflict of interest statement
Declaration of interests: The authors declare no competing interests.
Figures


References
-
- Artegiani B., Lyubimova A., Muraro M., van Es J. H., van Oudenaarden A., and Clevers H. (2017). A Single-Cell RNA Sequencing Study Reveals Cellular and Molecular Dynamics of the Hippocampal Neurogenic Niche. Cell Rep 21, 3271–3284. - PubMed
-
- Bonnay F., Veloso A., Steinmann V., Kocher T., Abdusselamoglu M. D., Bajaj S., Rivelles E., Landskron L., Esterbauer H., Zinzen R. P., and Knoblich J. A. (2020). Oxidative Metabolism Drives Immortalization of Neural Stem Cells during Tumorigenesis. Cell 182, 1490–1507 e1419. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous