

Dipeptide-Derived Alkynes as Potent and Selective Irreversible Inhibitors of Cysteine Cathepsins
- PMID: 36867428
- PMCID: PMC10041539
- DOI: 10.1021/acs.jmedchem.2c01360
Dipeptide-Derived Alkynes as Potent and Selective Irreversible Inhibitors of Cysteine Cathepsins
Abstract
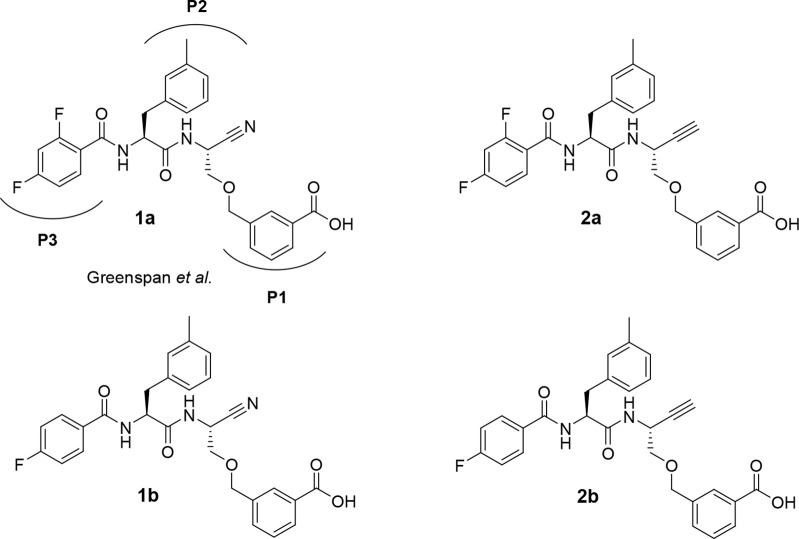
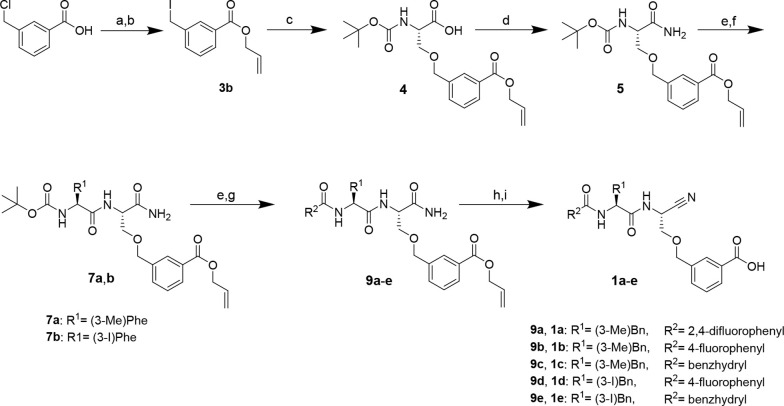
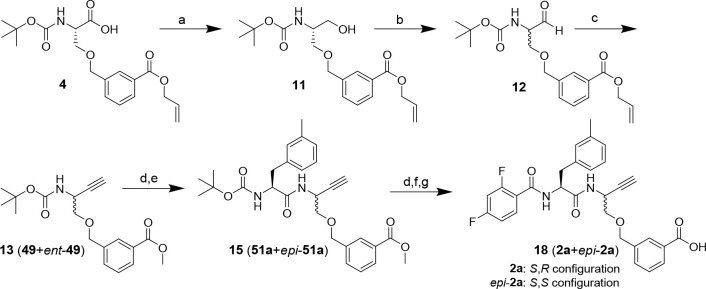
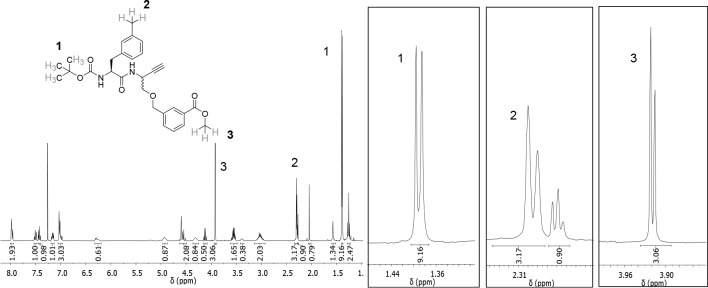
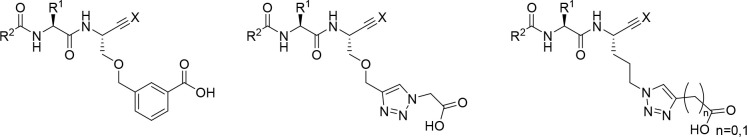
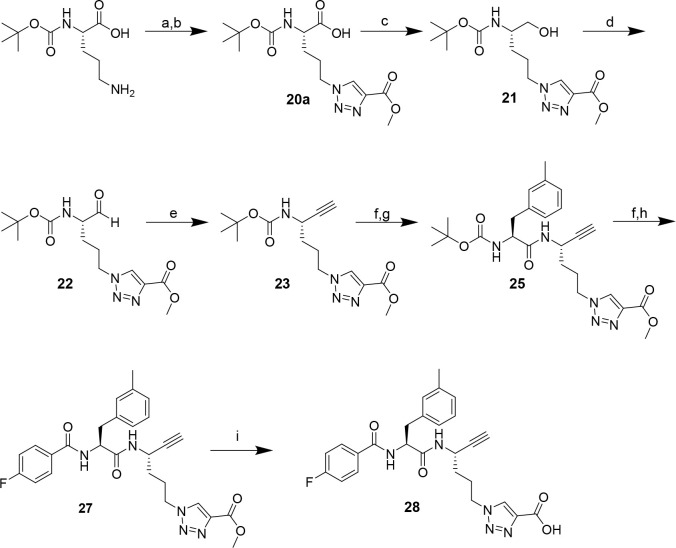
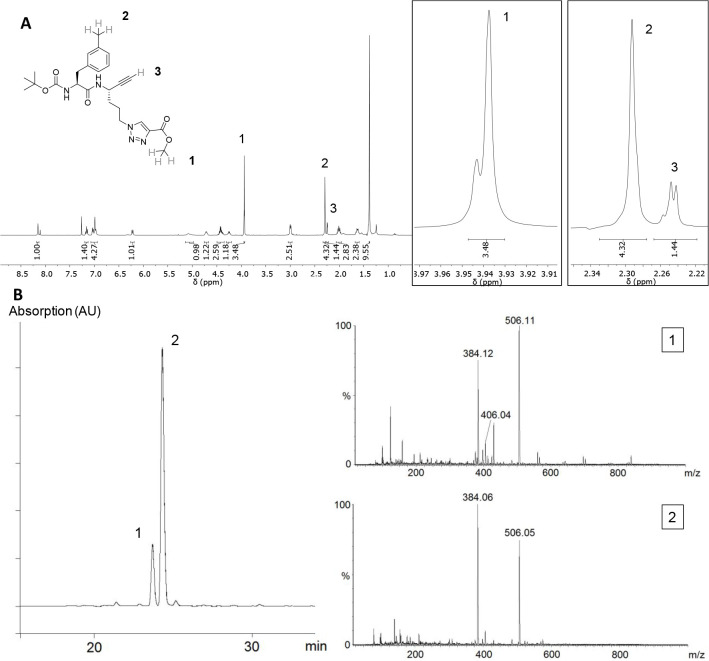
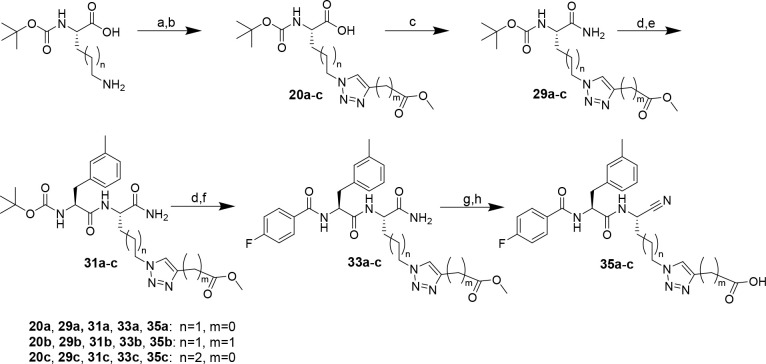
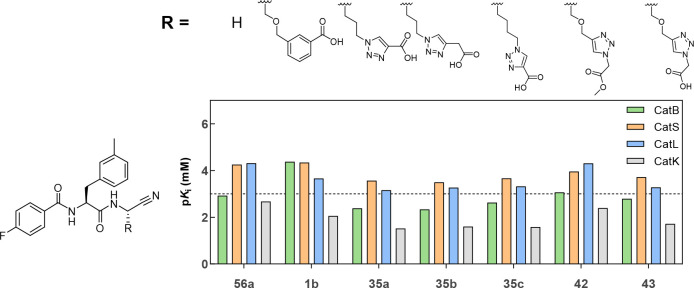
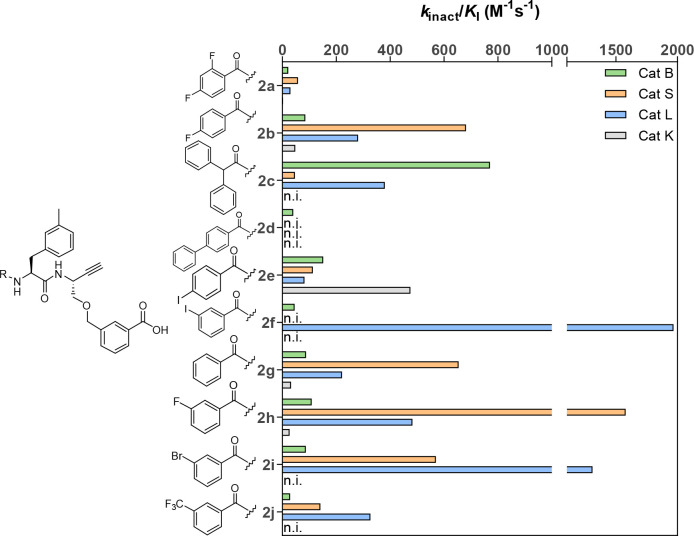
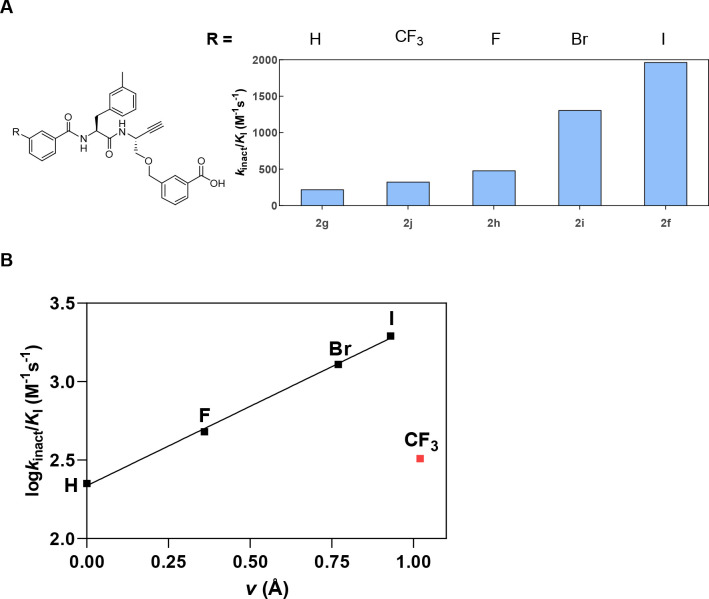
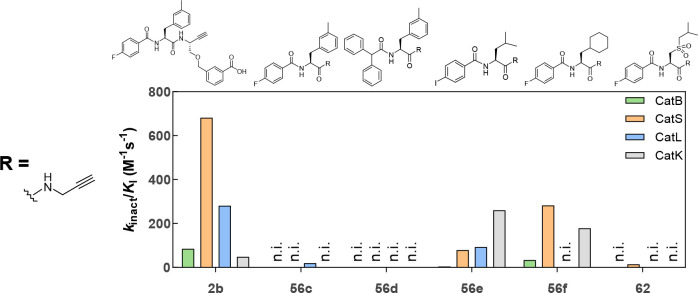
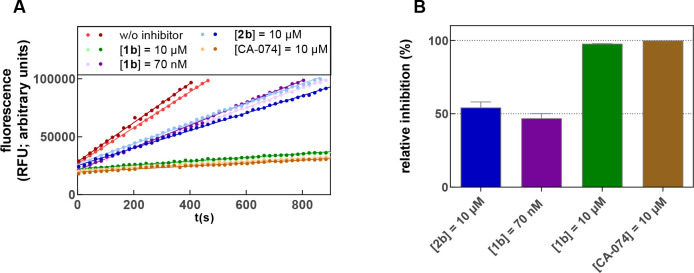
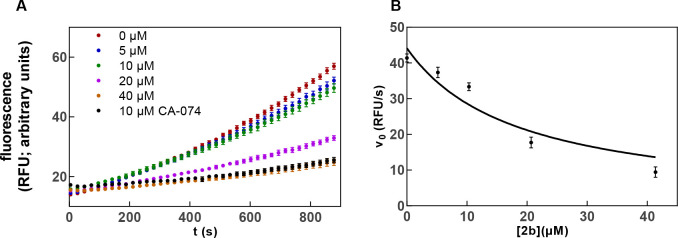
The potential of designing irreversible alkyne-based inhibitors of cysteine cathepsins by isoelectronic replacement in reversibly acting potent peptide nitriles was explored. The synthesis of the dipeptide alkynes was developed with special emphasis on stereochemically homogeneous products obtained in the Gilbert-Seyferth homologation for C≡C bond formation. Twenty-three dipeptide alkynes and 12 analogous nitriles were synthesized and investigated for their inhibition of cathepsins B, L, S, and K. Numerous combinations of residues at positions P1 and P2 as well as terminal acyl groups allowed for the derivation of extensive structure-activity relationships, which were rationalized by computational covalent docking for selected examples. The determined inactivation constants of the alkynes at the target enzymes span a range of >3 orders of magnitude (3-10 133 M-1 s-1). Notably, the selectivity profiles of alkynes do not necessarily reflect those of the nitriles. Inhibitory activity at the cellular level was demonstrated for selected compounds.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
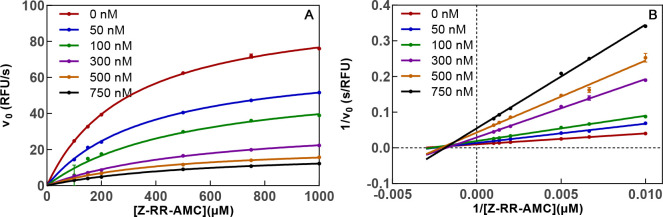
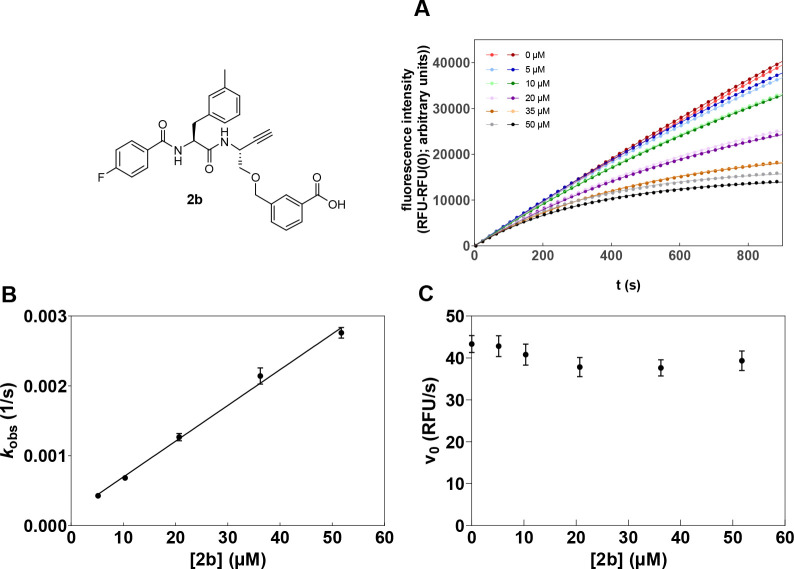
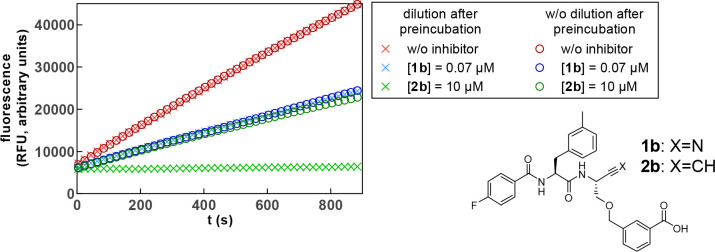
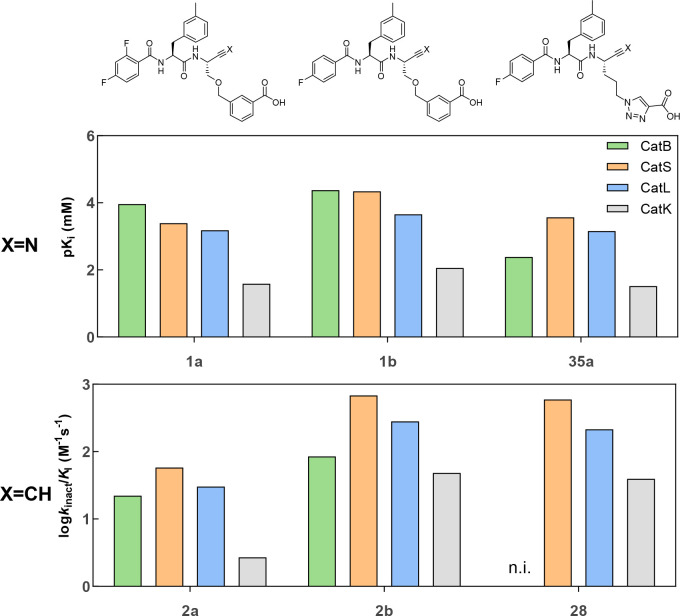
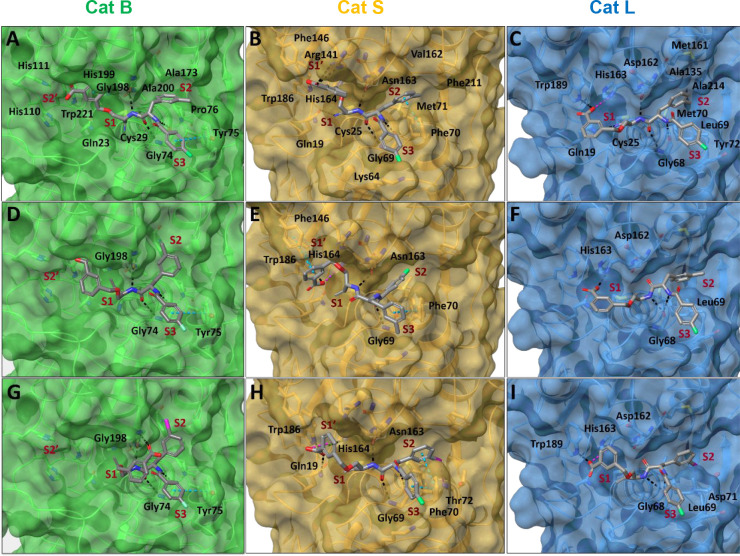
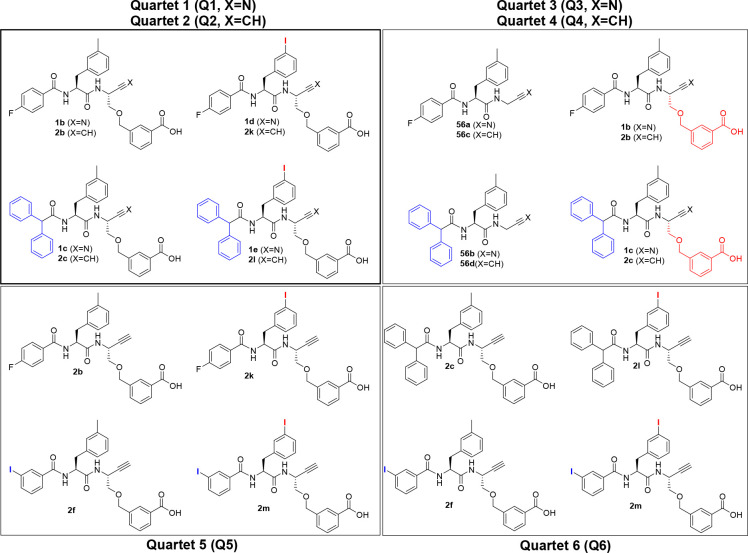
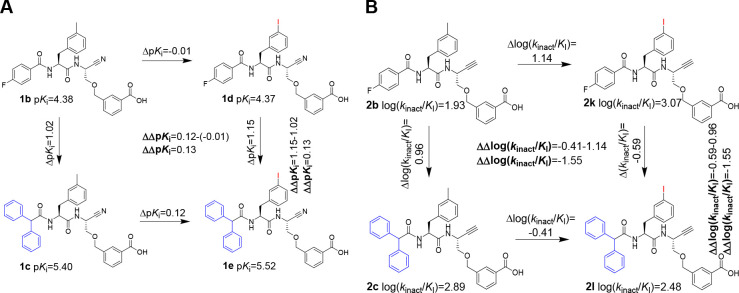
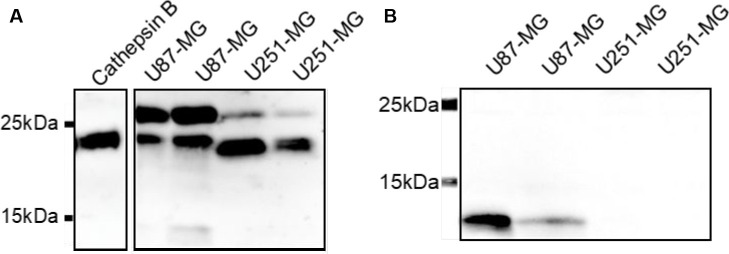
Similar articles
-
Dipeptide-derived nitriles containing additional electrophilic sites: potentially irreversible inhibitors of cysteine proteases.J Enzyme Inhib Med Chem. 2009 Dec;24(6):1245-52. doi: 10.3109/14756360902797328. J Enzyme Inhib Med Chem. 2009. PMID: 19912058
-
Fluorescent nitrile-based inhibitors of cysteine cathepsins.Bioorg Med Chem Lett. 2012 Dec 15;22(24):7715-8. doi: 10.1016/j.bmcl.2012.09.086. Epub 2012 Oct 3. Bioorg Med Chem Lett. 2012. PMID: 23122525
-
Active Site Mapping of Human Cathepsin F with Dipeptide Nitrile Inhibitors.ChemMedChem. 2015 Aug;10(8):1365-77. doi: 10.1002/cmdc.201500151. Epub 2015 Jun 26. ChemMedChem. 2015. PMID: 26119278
-
Development of nitrile-based peptidic inhibitors of cysteine cathepsins.Curr Top Med Chem. 2010;10(3):294-322. doi: 10.2174/156802610790725452. Curr Top Med Chem. 2010. PMID: 20166952 Review.
-
The consequences of lysosomotropism on the design of selective cathepsin K inhibitors.Chembiochem. 2006 Oct;7(10):1525-35. doi: 10.1002/cbic.200600149. Chembiochem. 2006. PMID: 16921579 Review.
Cited by
-
Development and Application of Reversible and Irreversible Covalent Probes for Human and Mouse Cathepsin-K Activity Detection, Revealing Nuclear Activity.Adv Sci (Weinh). 2024 Oct;11(38):e2401518. doi: 10.1002/advs.202401518. Epub 2024 Jul 5. Adv Sci (Weinh). 2024. PMID: 38970171 Free PMC article.
References
-
- Sloane B. F.; List K.; Fingleton B.; Matrisian L.. Proteases in Cancer: Significance for Invasion and Metastasis. In Proteases: Structure and Function; Brix K., Stöcker W., Eds.; Springer: Vienna, 2013; pp 491–550.
-
- Flores-Reséndiz D.; Castellanos-Juárez E.; Benítez-Bribiesca L. Las proteasas en la progresión neoplásica. Gac. Med. Mex. 2009, 145 (2), 131–142. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Chemical Information
