

Endothelial and Leptin Receptor+ cells promote the maintenance of stem cells and hematopoiesis in early postnatal murine bone marrow
- PMID: 36868235
- PMCID: PMC10035381
- DOI: 10.1016/j.devcel.2023.02.003
Endothelial and Leptin Receptor+ cells promote the maintenance of stem cells and hematopoiesis in early postnatal murine bone marrow
Abstract
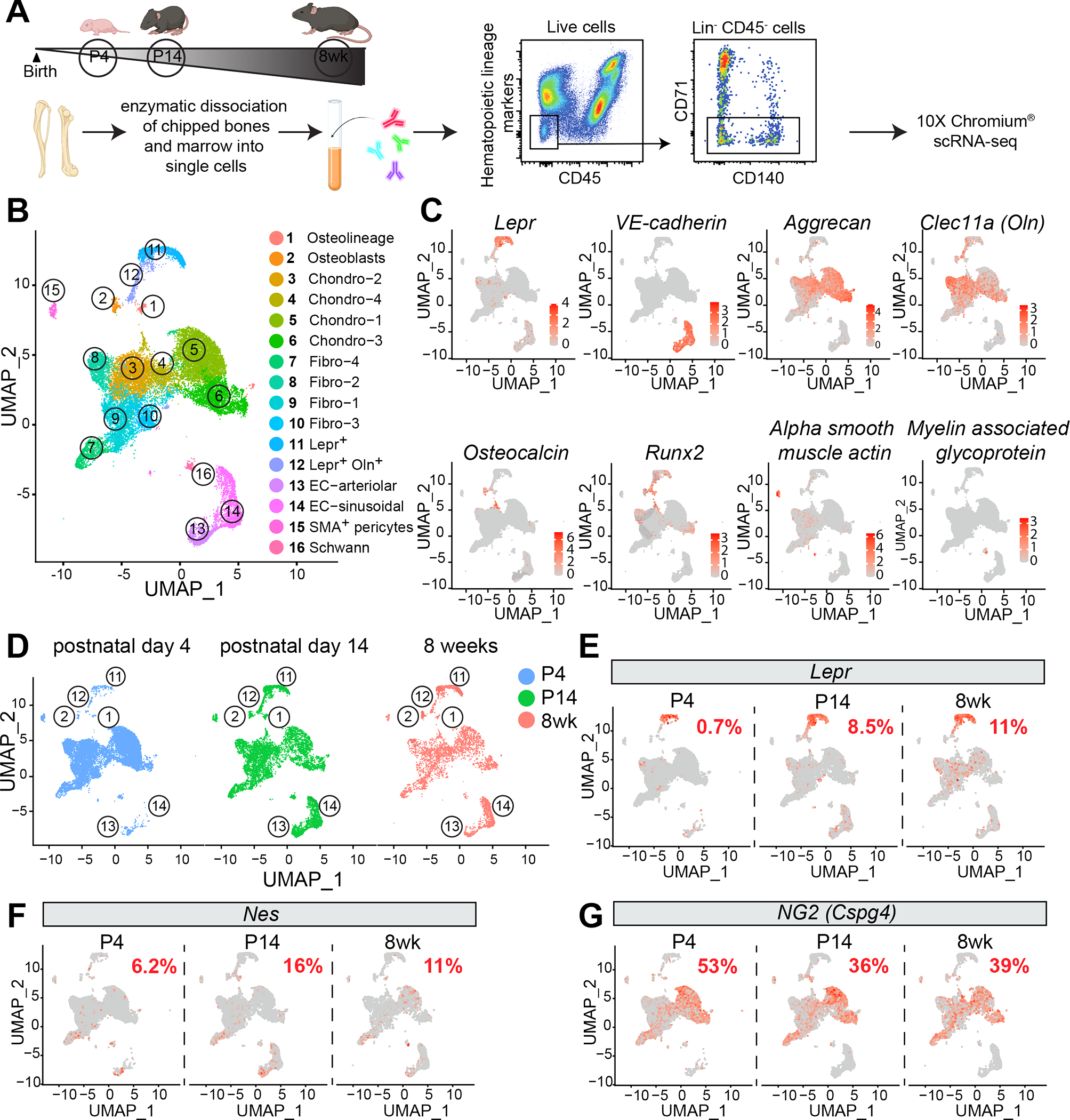
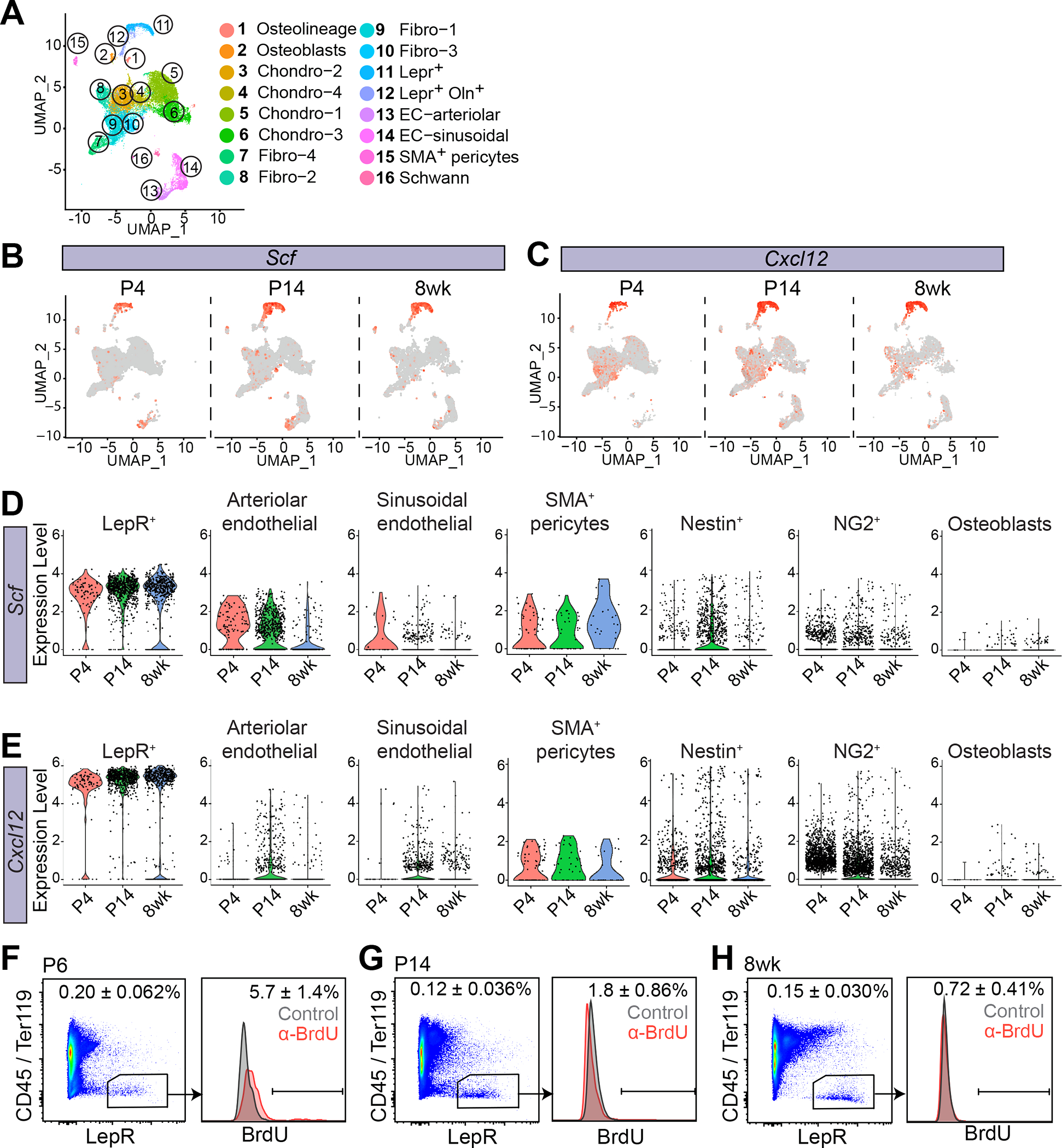
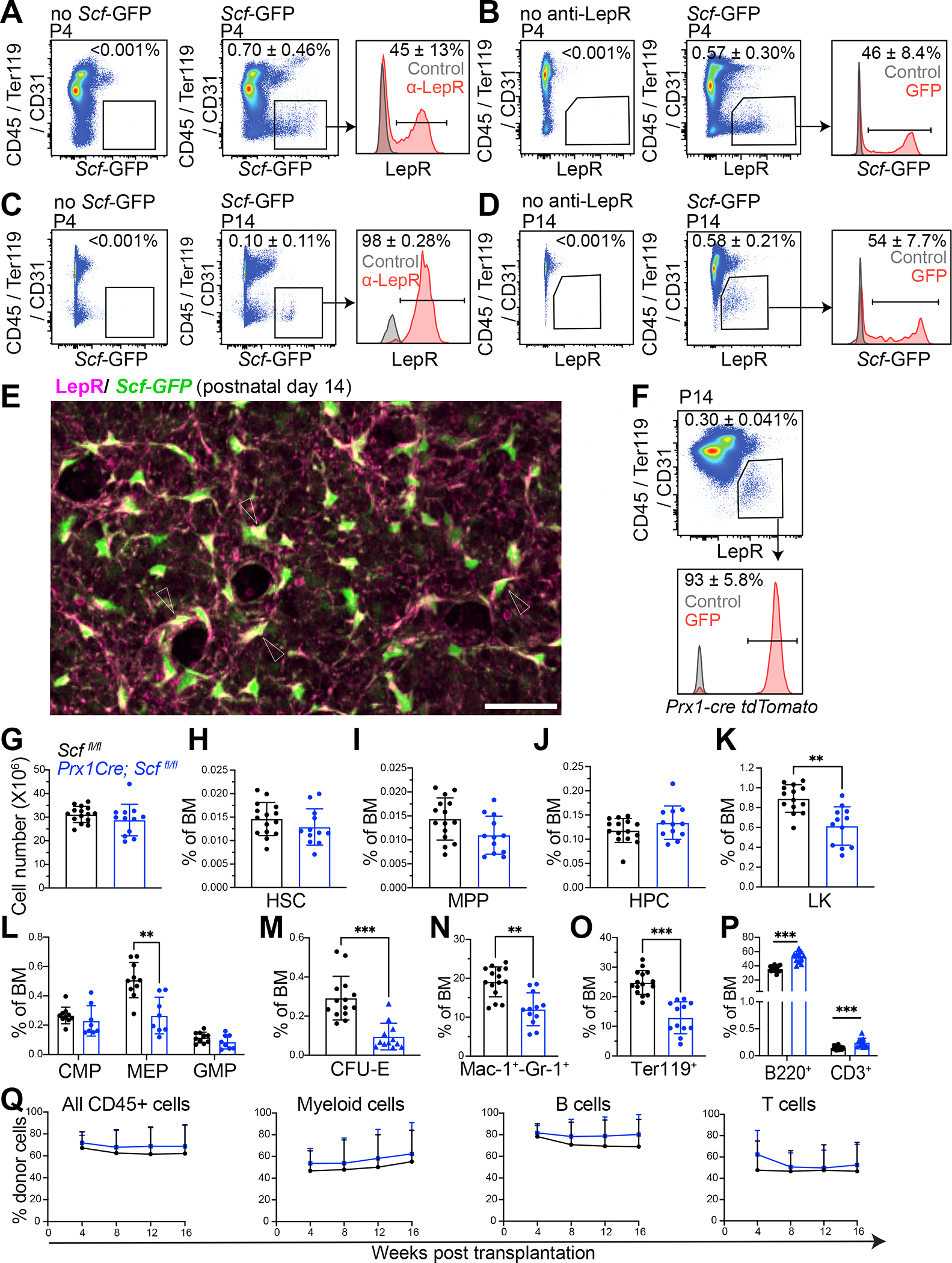
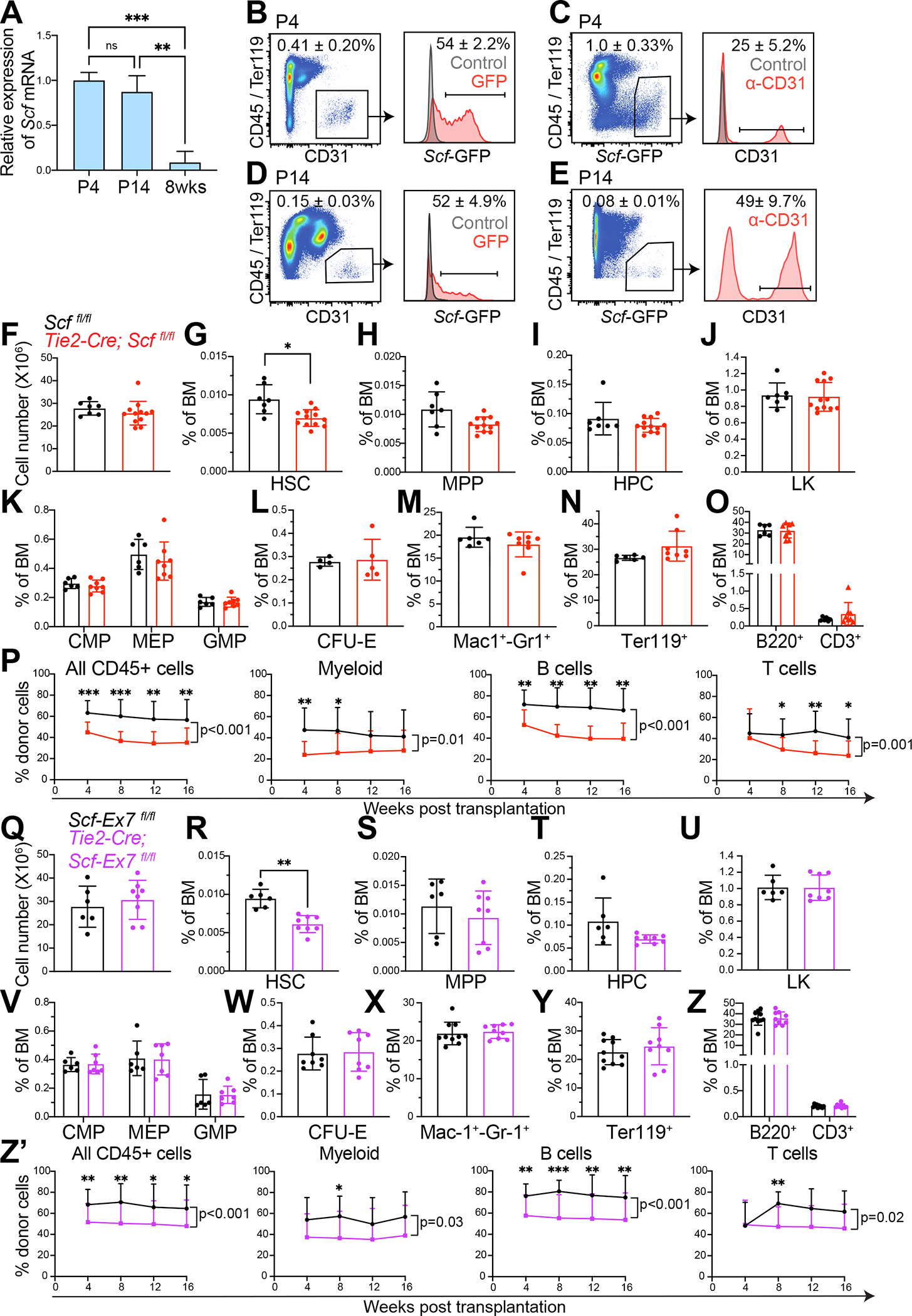
Mammalian hematopoietic stem cells (HSCs) colonize the bone marrow during late fetal development, and this becomes the major site of hematopoiesis after birth. However, little is known about the early postnatal bone marrow niche. We performed single-cell RNA sequencing of mouse bone marrow stromal cells at 4 days, 14 days, and 8 weeks after birth. Leptin-receptor-expressing (LepR+) stromal cells and endothelial cells increased in frequency during this period and changed their properties. At all postnatal stages, LepR+ cells and endothelial cells expressed the highest stem cell factor (Scf) levels in the bone marrow. LepR+ cells expressed the highest Cxcl12 levels. In early postnatal bone marrow, SCF from LepR+/Prx1+ stromal cells promoted myeloid and erythroid progenitor maintenance, while SCF from endothelial cells promoted HSC maintenance. Membrane-bound SCF in endothelial cells contributed to HSC maintenance. LepR+ cells and endothelial cells are thus important niche components in early postnatal bone marrow.
Keywords: erythropoiesis; myelopoiesis; niche; stem cell factor; stromal cells.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
