

Genetic requirements for repair of lesions caused by single genomic ribonucleotides in S phase
- PMID: 36869098
- PMCID: PMC9984532
- DOI: 10.1038/s41467-023-36866-6
Genetic requirements for repair of lesions caused by single genomic ribonucleotides in S phase
Abstract
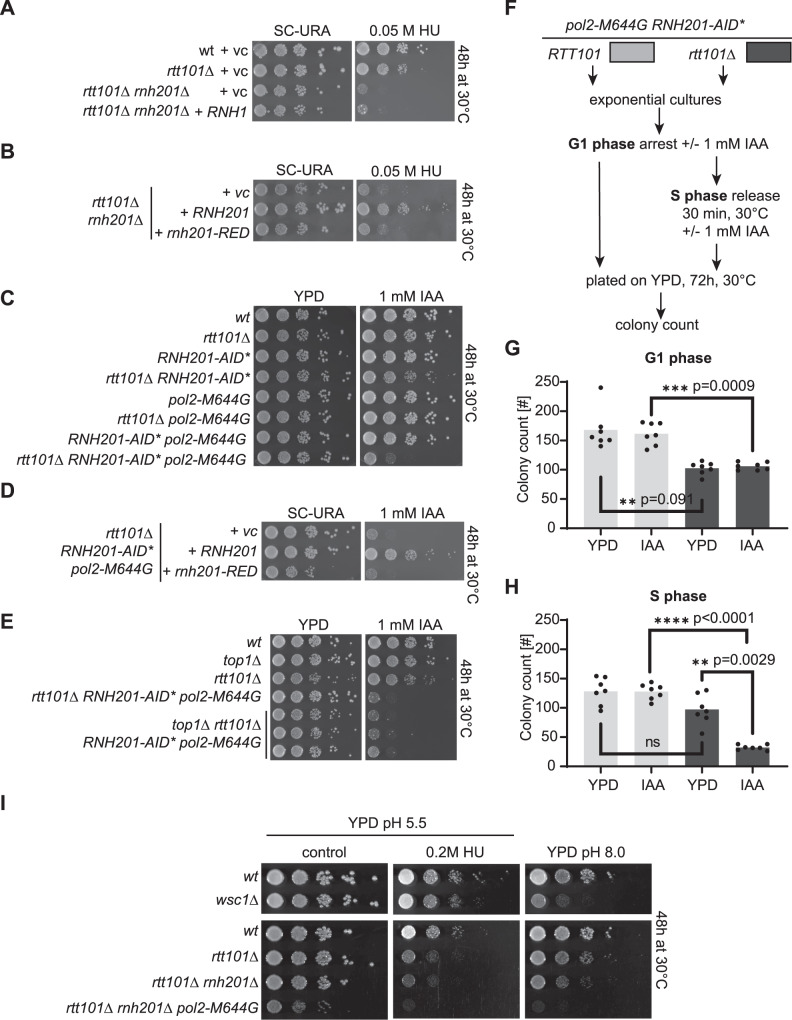
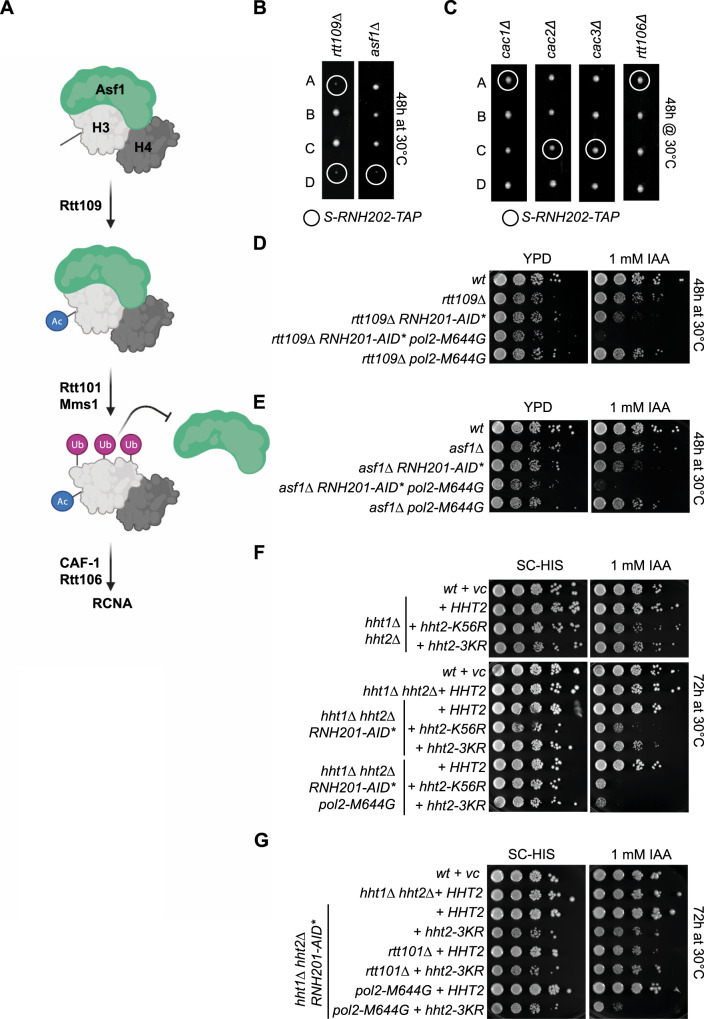
Single ribonucleoside monophosphates (rNMPs) are transiently present in eukaryotic genomes. The RNase H2-dependent ribonucleotide excision repair (RER) pathway ensures error-free rNMP removal. In some pathological conditions, rNMP removal is impaired. If these rNMPs hydrolyze during, or prior to, S phase, toxic single-ended double-strand breaks (seDSBs) can occur upon an encounter with replication forks. How such rNMP-derived seDSB lesions are repaired is unclear. We expressed a cell cycle phase restricted allele of RNase H2 to nick at rNMPs in S phase and study their repair. Although Top1 is dispensable, the RAD52 epistasis group and Rtt101Mms1-Mms22 dependent ubiquitylation of histone H3 become essential for rNMP-derived lesion tolerance. Consistently, loss of Rtt101Mms1-Mms22 combined with RNase H2 dysfunction leads to compromised cellular fitness. We refer to this repair pathway as nick lesion repair (NLR). The NLR genetic network may have important implications in the context of human pathologies.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
