

Hello darkness, my old friend: 3-KETOACYL-COENZYME A SYNTHASE4 is a branch point in the regulation of triacylglycerol synthesis in Arabidopsis thaliana
- PMID: 36869652
- PMCID: PMC10226560
- DOI: 10.1093/plcell/koad059
Hello darkness, my old friend: 3-KETOACYL-COENZYME A SYNTHASE4 is a branch point in the regulation of triacylglycerol synthesis in Arabidopsis thaliana
Abstract
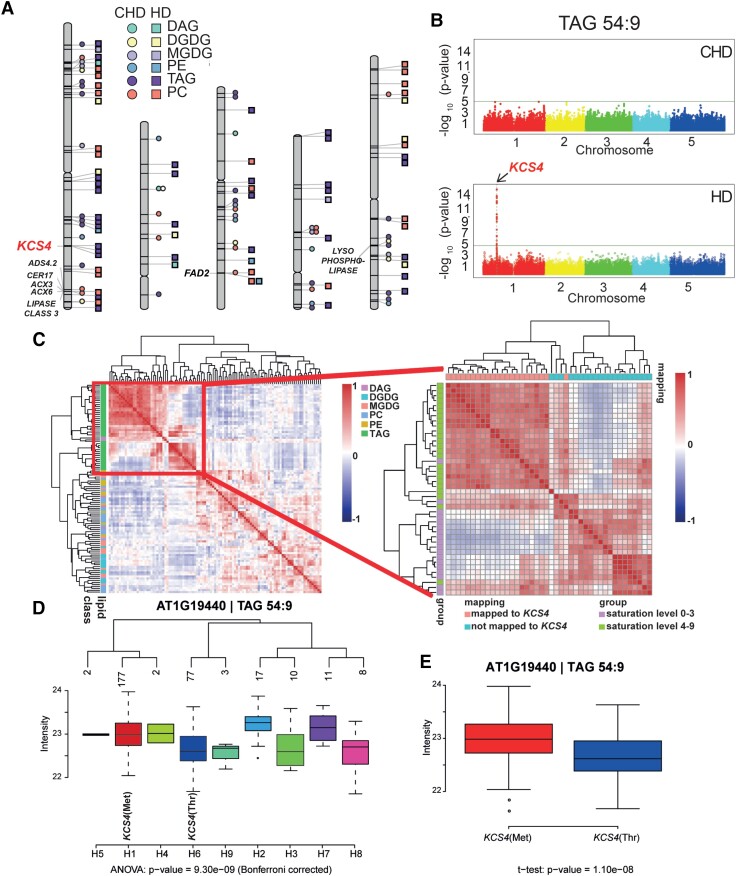
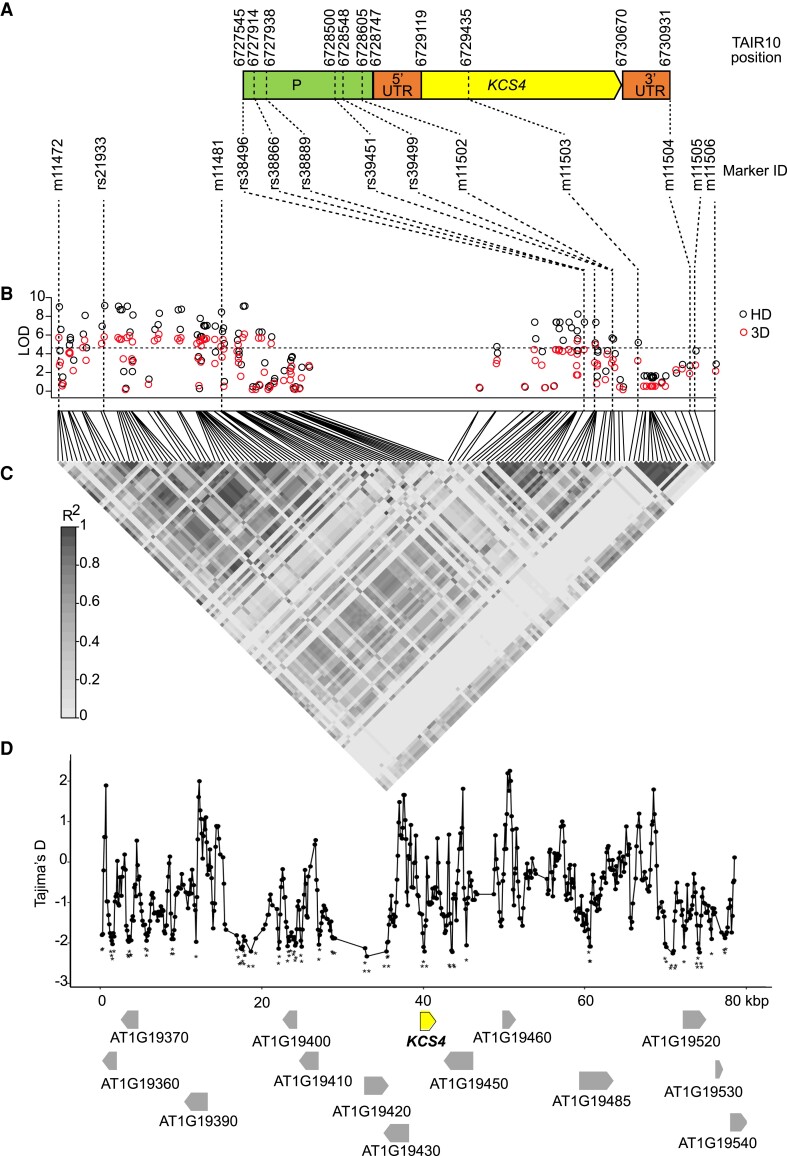
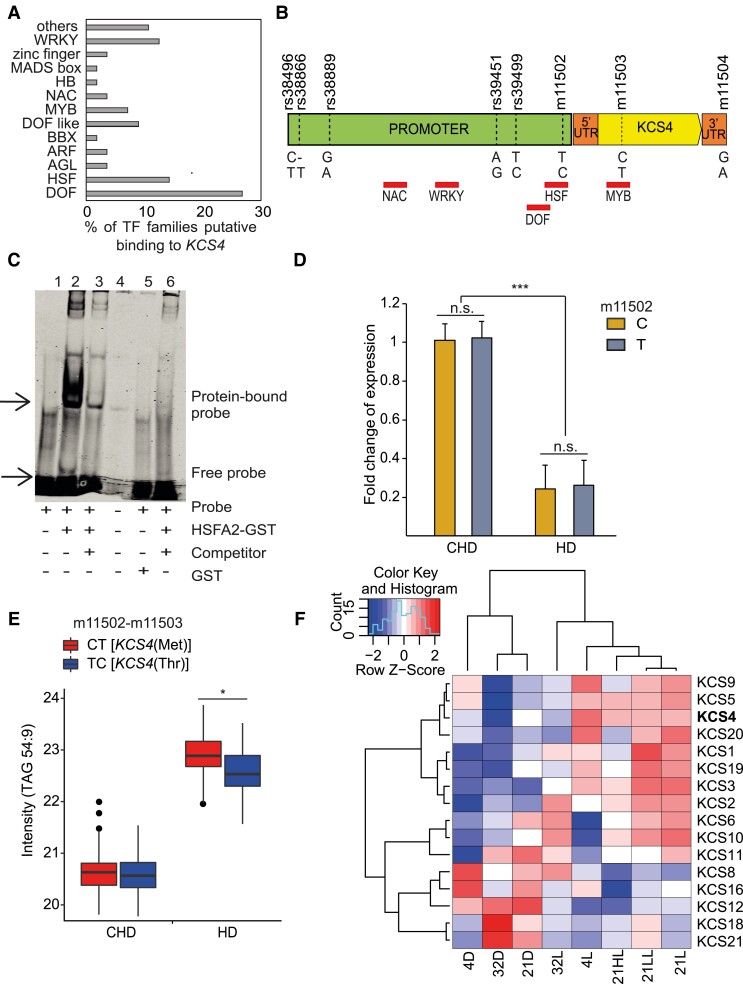
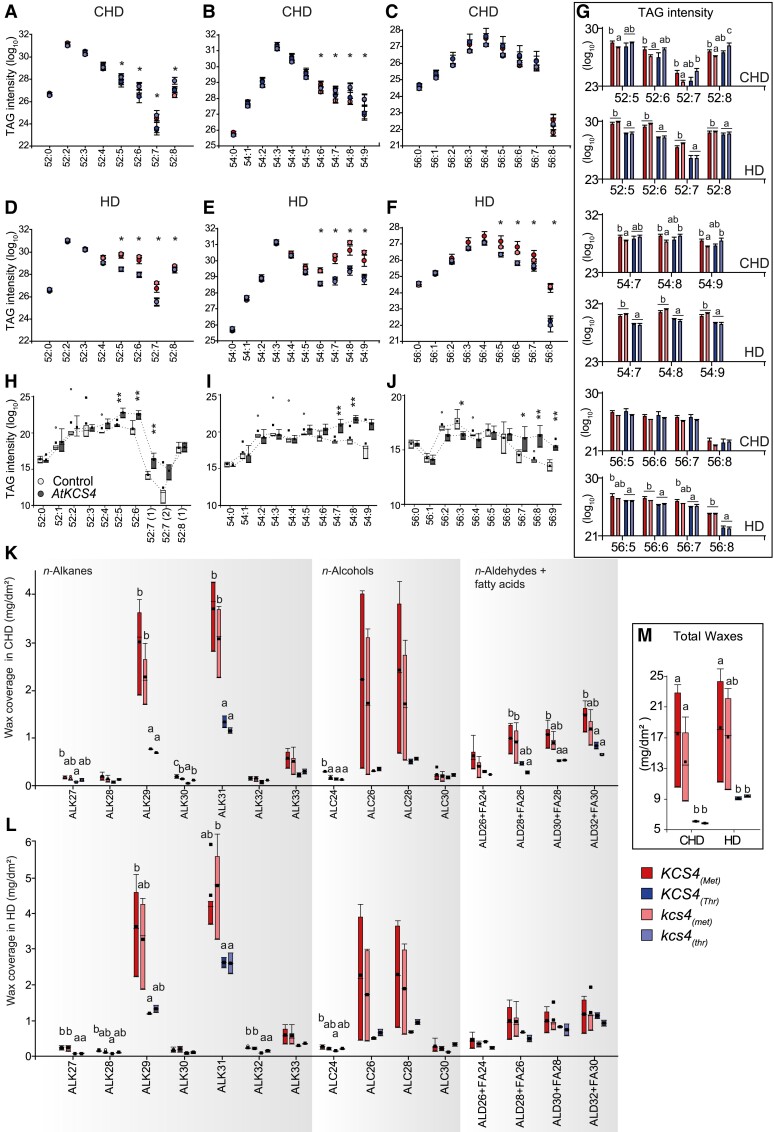
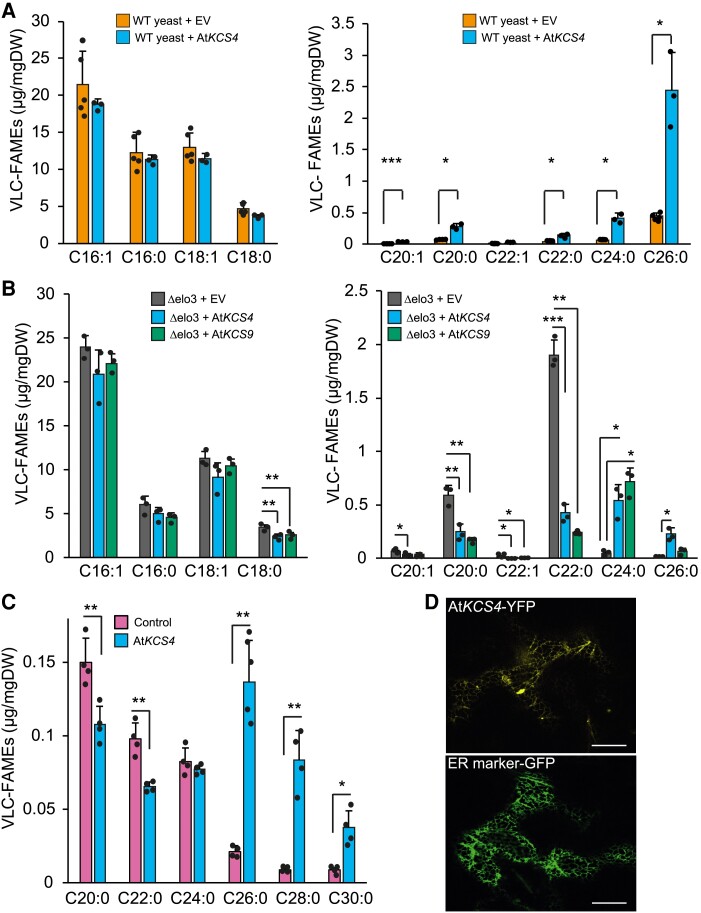
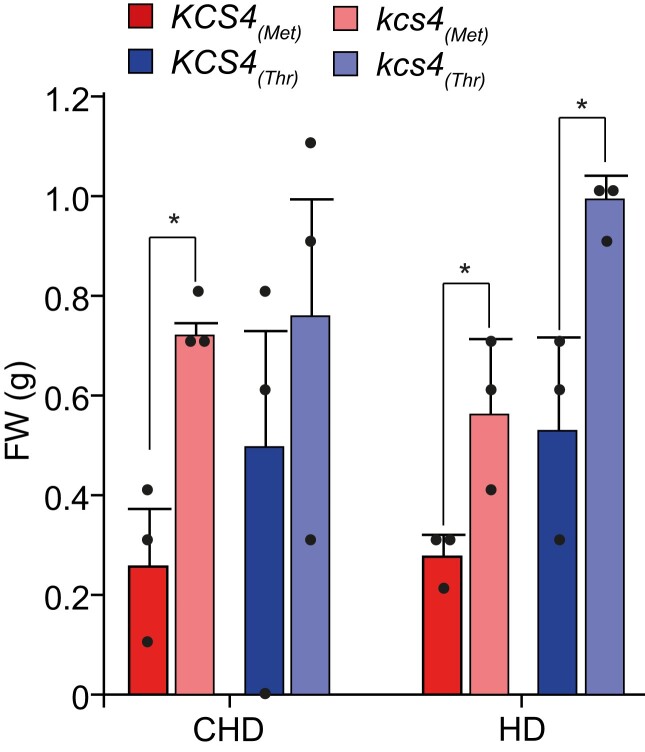
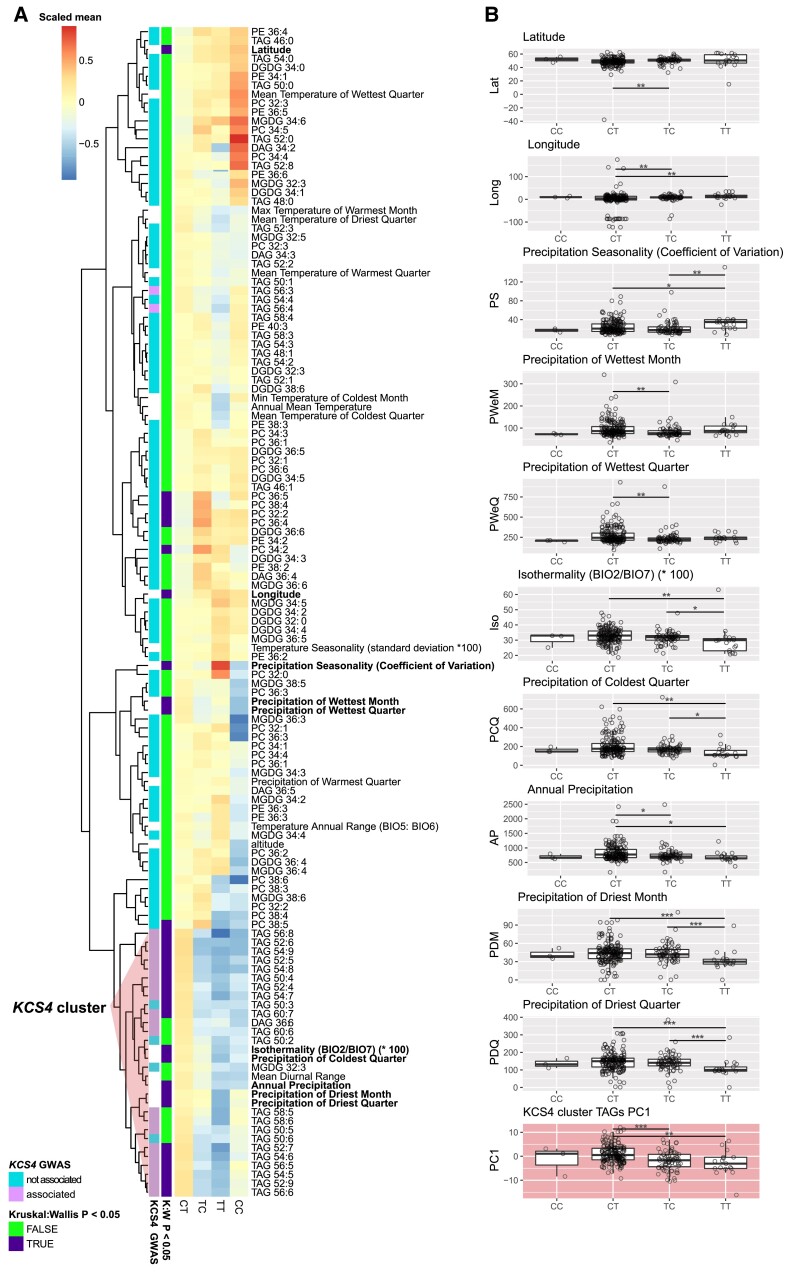
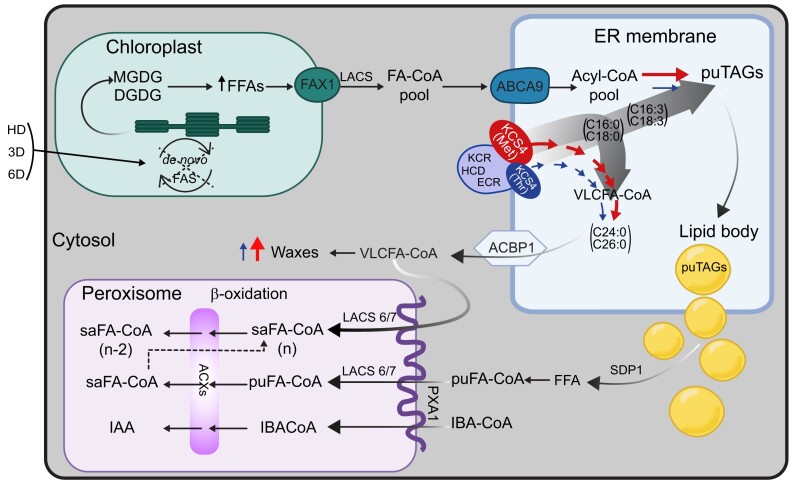
Plant lipids are important as alternative sources of carbon and energy when sugars or starch are limited. Here, we applied combined heat and darkness or extended darkness to a panel of ∼300 Arabidopsis (Arabidopsis thaliana) accessions to study lipid remodeling under carbon starvation. Natural allelic variation at 3-KETOACYL-COENZYME A SYNTHASE4 (KCS4), a gene encoding an enzyme involved in very long chain fatty acid (VLCFA) synthesis, underlies the differential accumulation of polyunsaturated triacylglycerols (puTAGs) under stress. Ectopic expression of KCS4 in yeast and plants proved that KCS4 is a functional enzyme localized in the endoplasmic reticulum with specificity for C22 and C24 saturated acyl-CoA. Allelic mutants and transient overexpression in planta revealed the differential role of KCS4 alleles in VLCFA synthesis and leaf wax coverage, puTAG accumulation, and biomass. Moreover, the region harboring KCS4 is under high selective pressure and allelic variation at KCS4 correlates with environmental parameters from the locales of Arabidopsis accessions. Our results provide evidence that KCS4 plays a decisive role in the subsequent fate of fatty acids released from chloroplast membrane lipids under carbon starvation. This work sheds light on both plant response mechanisms and the evolutionary events shaping the lipidome under carbon starvation.
© The Author(s) 2023. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures
References
-
- Anderson MJ. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001:26(1):32–46. 10.1111/j.1442-9993.2001.01070.pp.x - DOI
-
- Baxter I, Brazelton JN, Yu D, Huang YS, Lahner B, Yakubova E, Li Y, Bergelson J, Borevitz JO, Nordborg M, et al. A coastal cline in sodium accumulation in Arabidopsis thaliana is driven by natural variation of the sodium transporter AtHKT1; 1. PLoS Genet. 2010:6(11):e1001193. 10.1371/journal.pgen.1001193 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
