

Stiffness Restricts the Stemness of the Intestinal Stem Cells and Skews Their Differentiation Toward Goblet Cells
- PMID: 36871599
- PMCID: PMC10200762
- DOI: 10.1053/j.gastro.2023.02.030
Stiffness Restricts the Stemness of the Intestinal Stem Cells and Skews Their Differentiation Toward Goblet Cells
Abstract
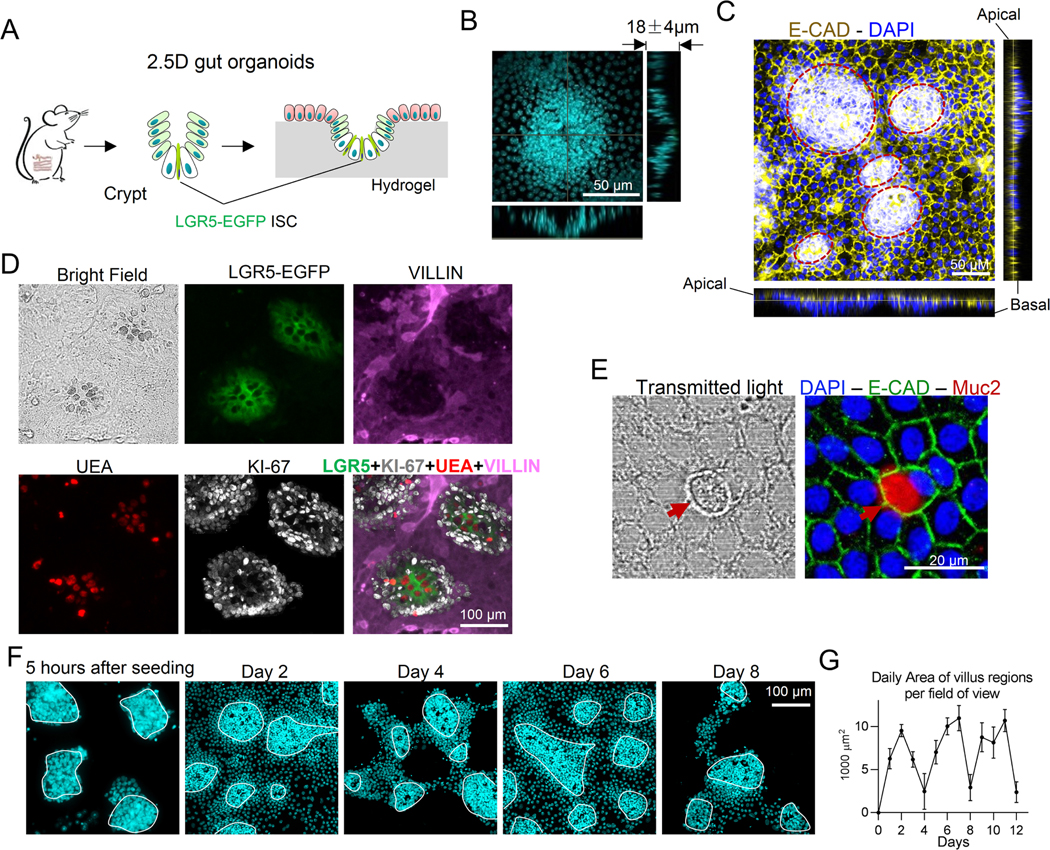
Background & aims: Fibrosis and tissue stiffening are hallmarks of inflammatory bowel disease (IBD). We have hypothesized that the increased stiffness directly contributes to the dysregulation of the epithelial cell homeostasis in IBD. Here, we aim to determine the impact of tissue stiffening on the fate and function of the intestinal stem cells (ISCs).
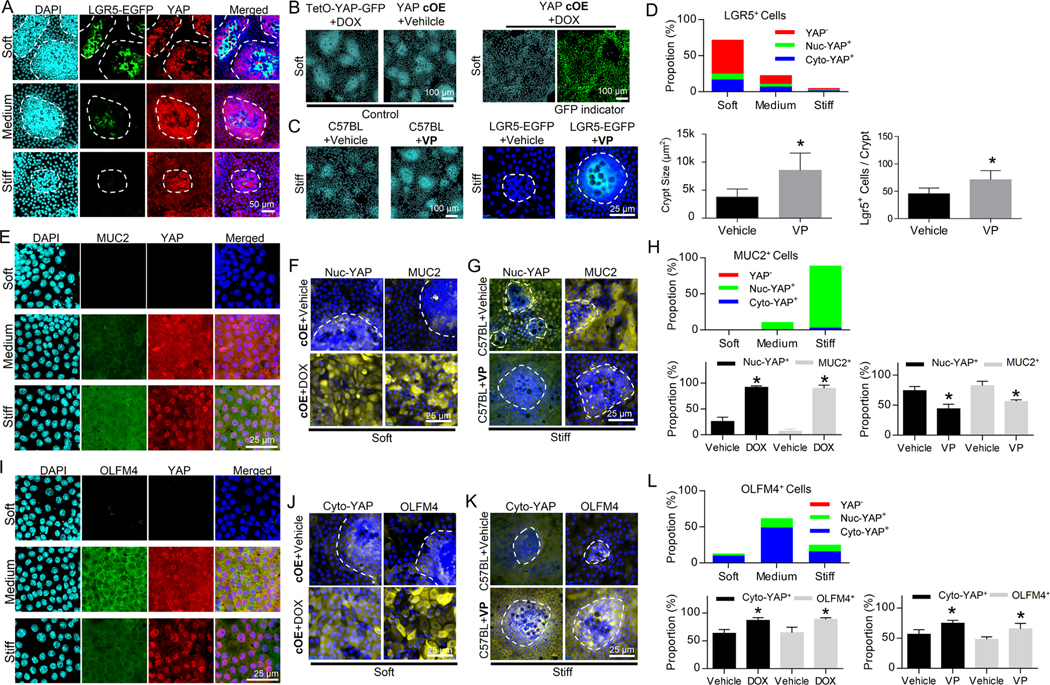
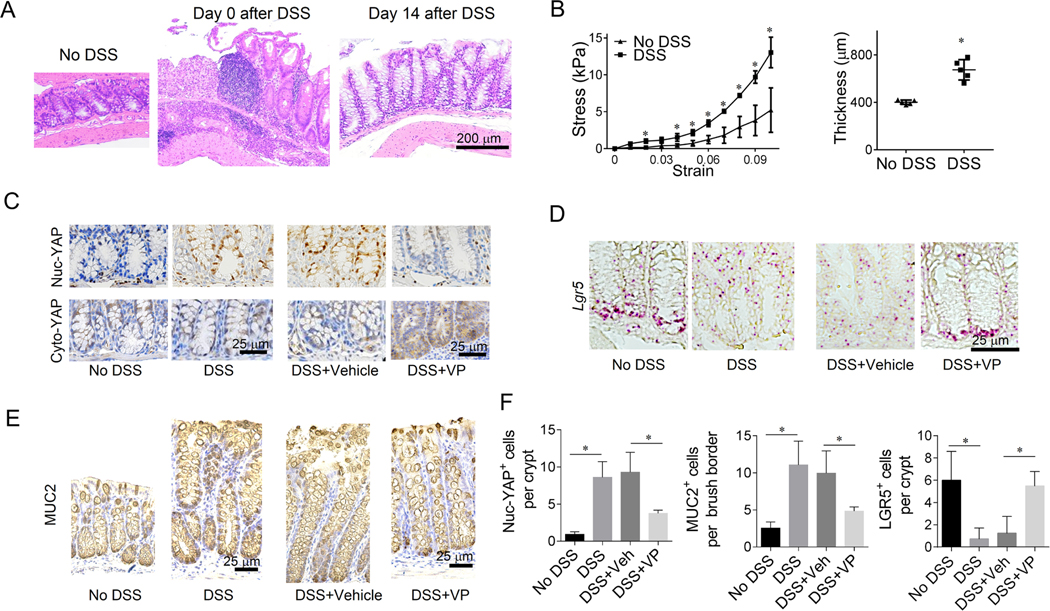
Methods: We developed a long-term culture system consisting of 2.5-dimensional intestinal organoids grown on a hydrogel matrix with tunable stiffness. Single-cell RNA sequencing provided stiffness-regulated transcriptional signatures of the ISCs and their differentiated progeny. YAP-knockout and YAP-overexpression mice were used to manipulate YAP expression. In addition, we analyzed colon samples from murine colitis models and human IBD samples to assess the impact of stiffness on ISCs in vivo.
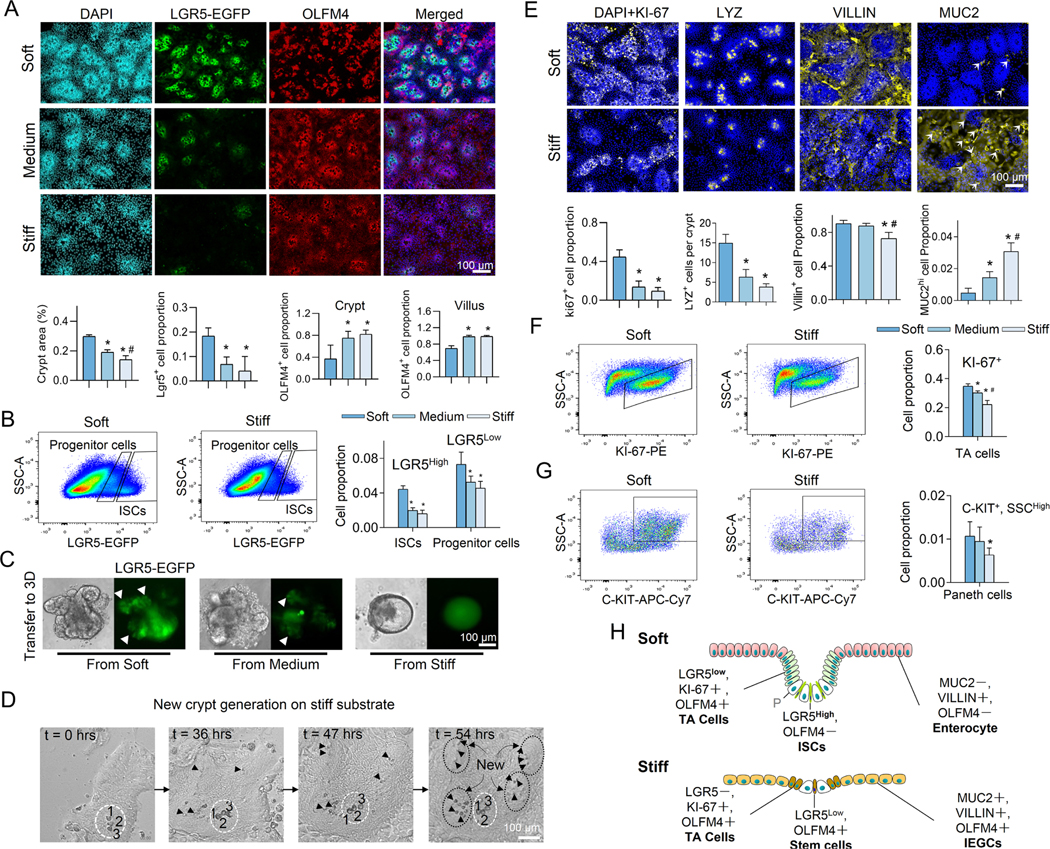
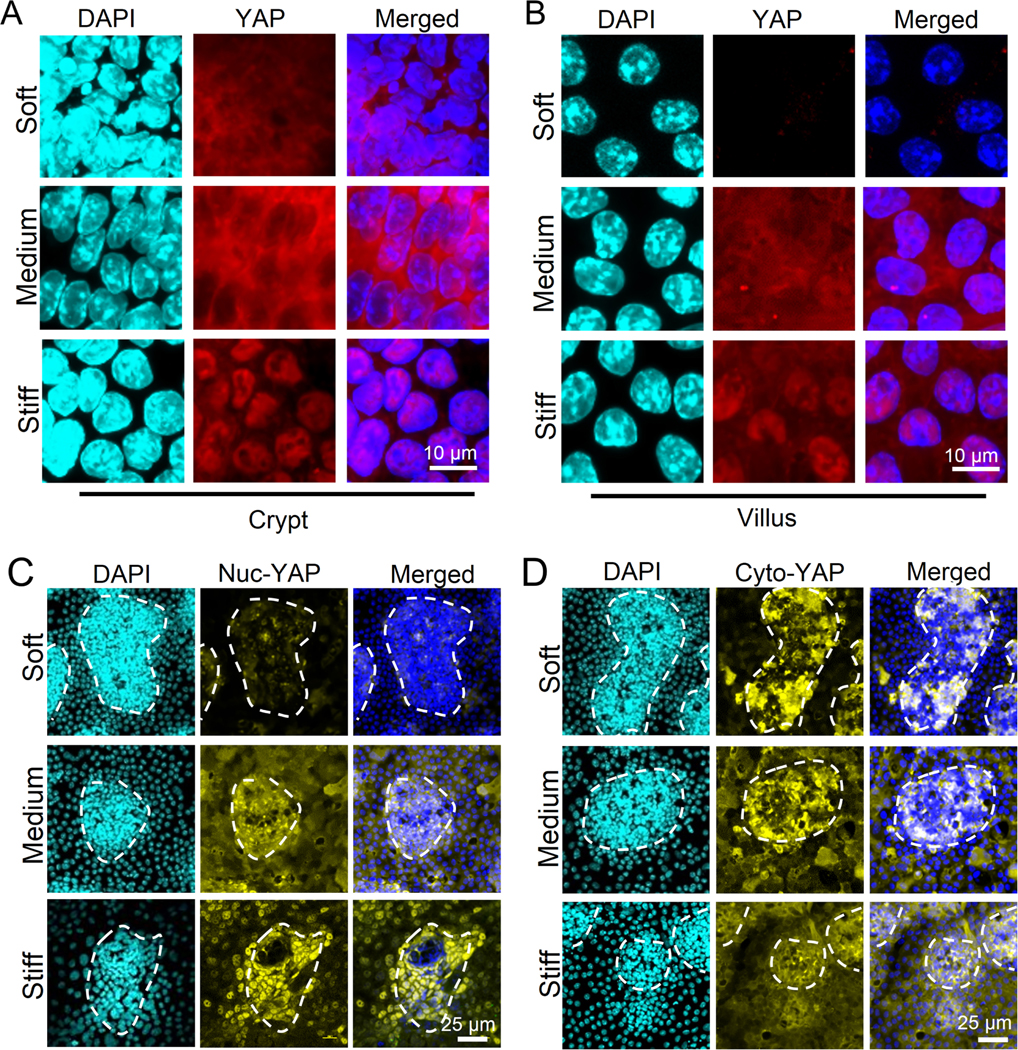
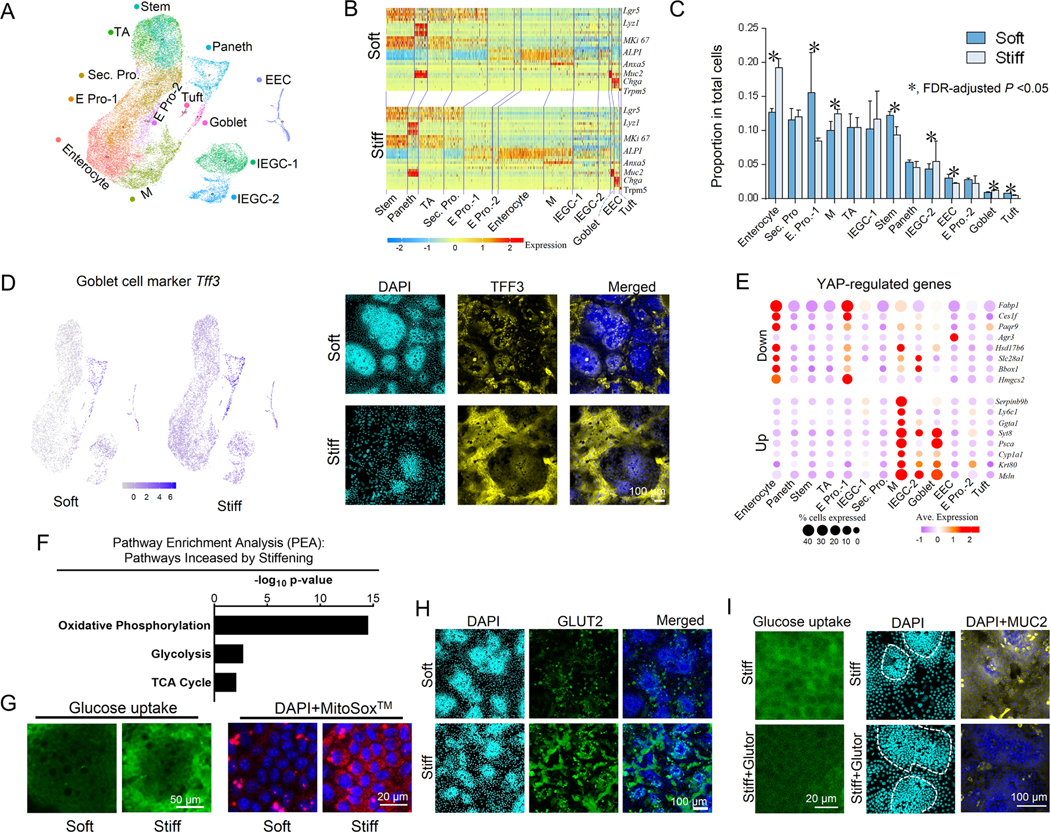
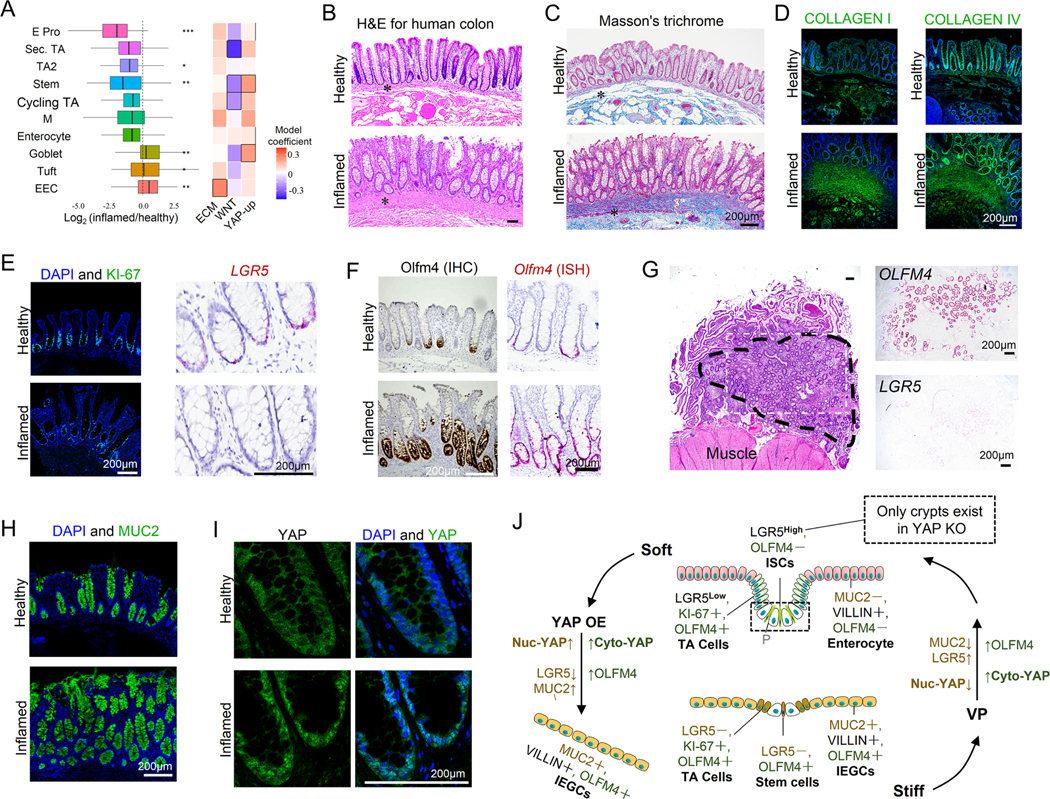
Results: We demonstrated that increasing the stiffness potently reduced the population of LGR5+ ISCs and KI-67+-proliferating cells. Conversely, cells expressing the stem cell marker, olfactomedin-4, became dominant in the crypt-like compartments and pervaded the villus-like regions. Concomitantly, stiffening prompted the ISCs to preferentially differentiate toward goblet cells. Mechanistically, stiffening increased the expression of cytosolic YAP, driving the extension of olfactomedin-4+ cells into the villus-like regions, while it induced the nuclear translocation of YAP, leading to preferential differentiation of ISCs toward goblet cells. Furthermore, analysis of colon samples from murine colitis models and patients with IBD demonstrated cellular and molecular remodeling reminiscent of those observed in vitro.
Conclusions: Collectively, our findings highlight that matrix stiffness potently regulates the stemness of ISCs and their differentiation trajectory, supporting the hypothesis that fibrosis-induced gut stiffening plays a direct role in epithelial remodeling in IBD.
Keywords: Fibrosis; IBD; Intestinal Organoids; Intestinal Stem Cells; Stiffening.
Copyright © 2023 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Bloemendaal AL, Buchs NC, George BD, et al. Intestinal stem cells and intestinal homeostasis in health and in inflammation: a review. Surgery 2016;159:1237–1248. - PubMed
-
- Schmitt M, Schewe M, Sacchetti A, et al. Paneth cells respond to inflammation and contribute to tissue regeneration by acquiring stem-like features through SCF/c-Kit signaling. Cell reports 2018;24:2312–2328. e7. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
