

Accessory ESCRT-III proteins are conserved and selective regulators of Rab11a-exosome formation
- PMID: 36872252
- PMCID: PMC9986085
- DOI: 10.1002/jev2.12311
Accessory ESCRT-III proteins are conserved and selective regulators of Rab11a-exosome formation
Erratum in
-
Correction to "Accessory ESCRT-III proteins are conserved and selective regulators of Rab11a-exosome formation".J Extracell Vesicles. 2023 May;12(5):e12321. doi: 10.1002/jev2.12321. J Extracell Vesicles. 2023. PMID: 37132054 Free PMC article. No abstract available.
Abstract
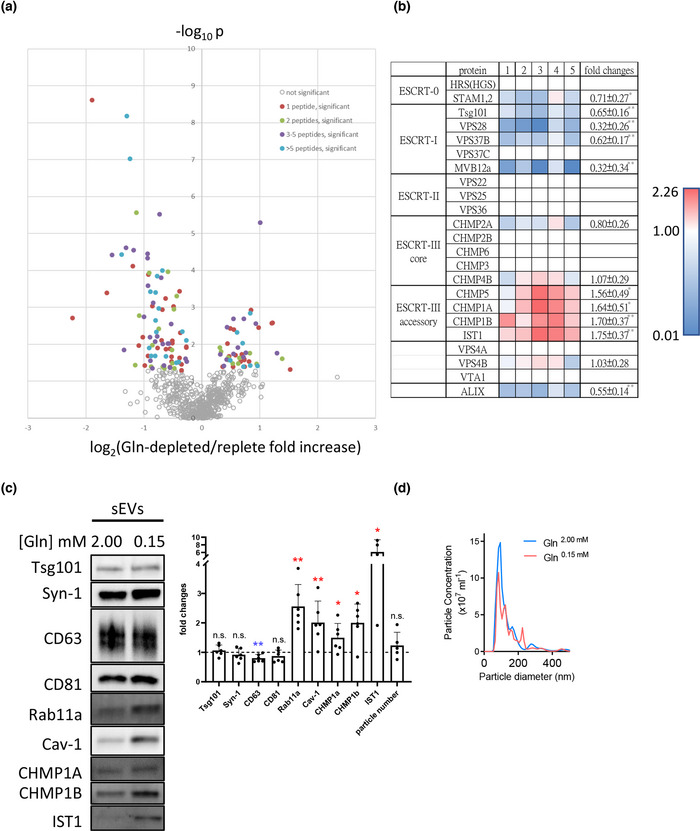
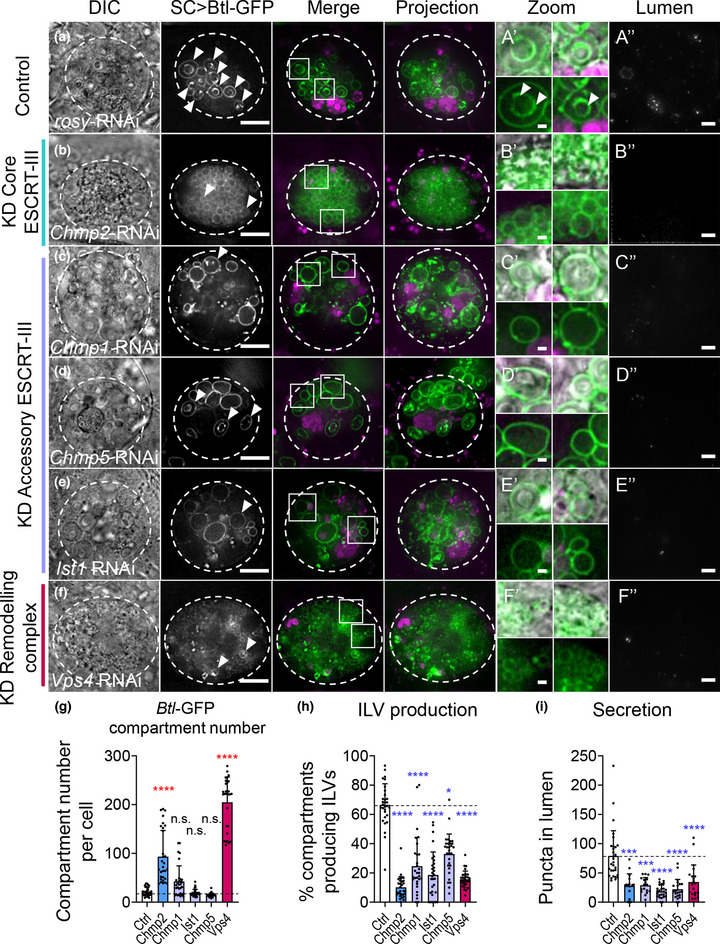
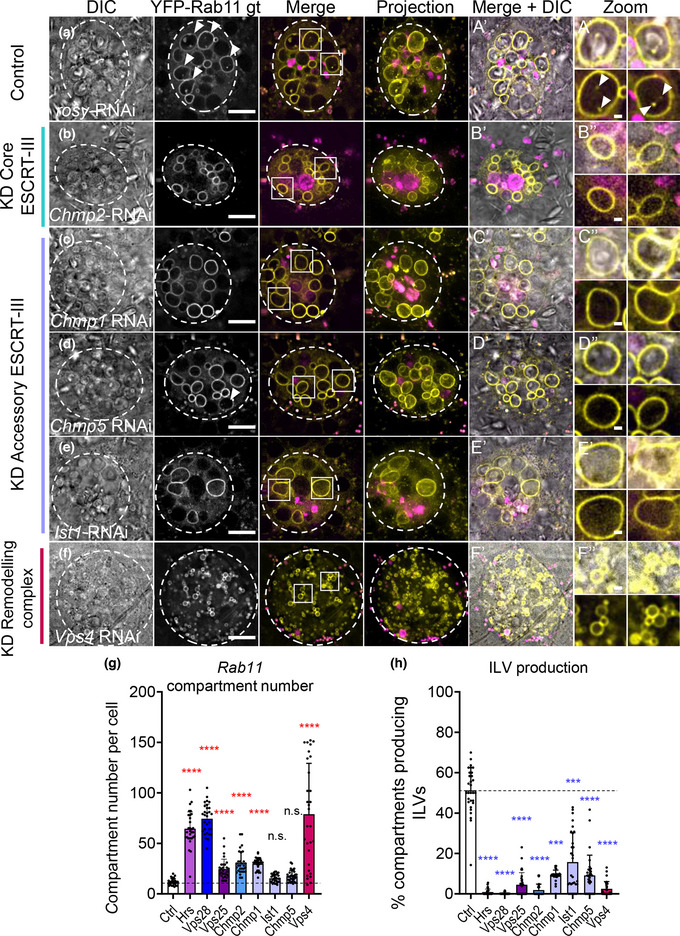
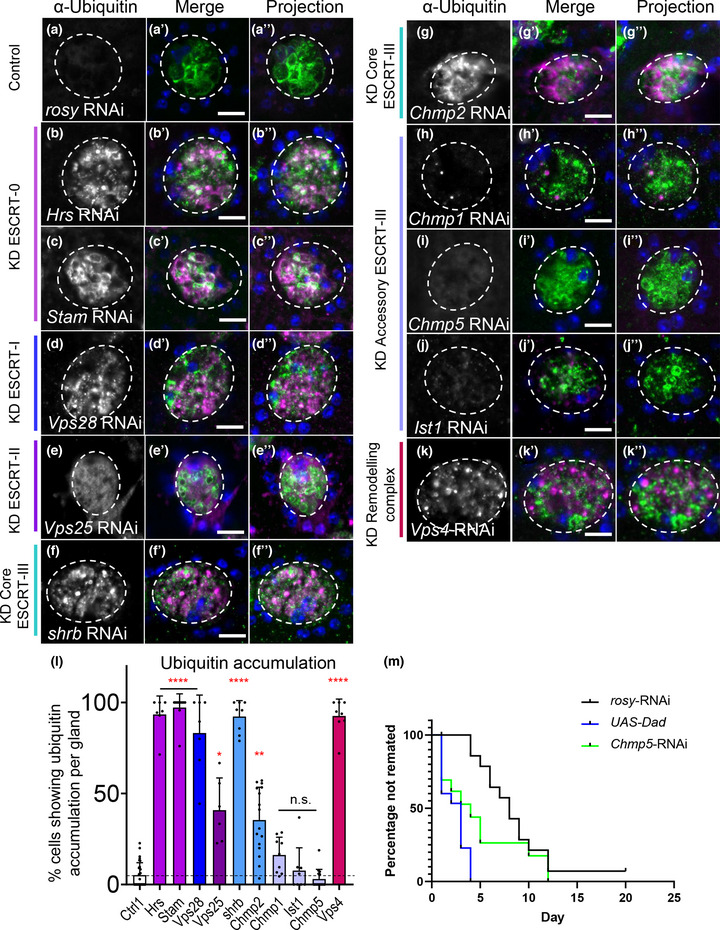
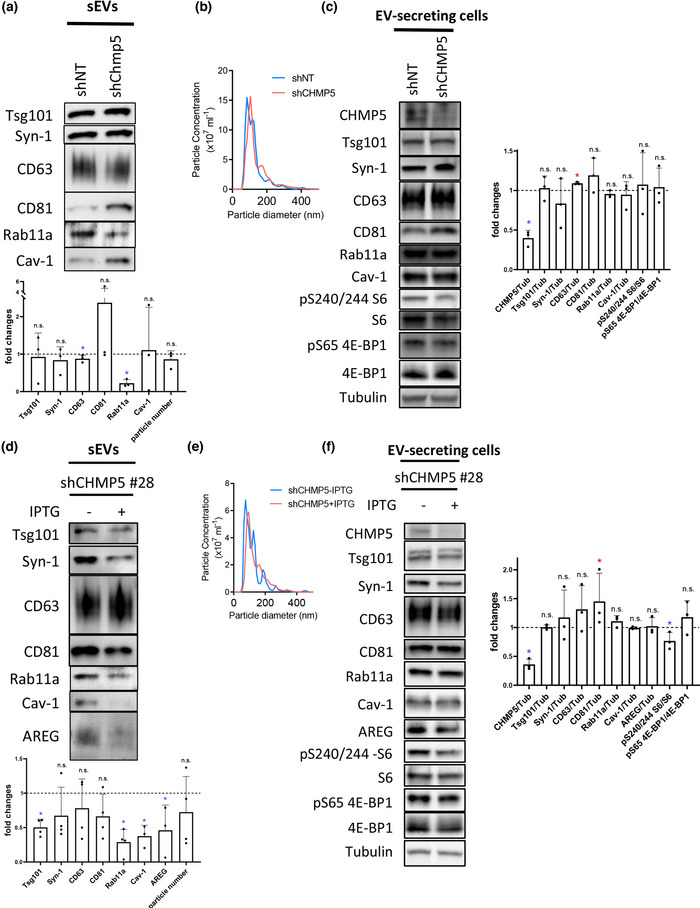
Exosomes are secreted nanovesicles with potent signalling activity that are initially formed as intraluminal vesicles (ILVs) in late Rab7-positive multivesicular endosomes, and also in recycling Rab11a-positive endosomes, particularly under some forms of nutrient stress. The core proteins of the Endosomal Sorting Complex Required for Transport (ESCRT) participate in exosome biogenesis and ILV-mediated destruction of ubiquitinylated cargos. Accessory ESCRT-III components have reported roles in ESCRT-III-mediated vesicle scission, but their precise functions are poorly defined. They frequently only appear essential under stress. Comparative proteomics analysis of human small extracellular vesicles revealed that accessory ESCRT-III proteins, CHMP1A, CHMP1B, CHMP5 and IST1, are increased in Rab11a-enriched exosome preparations. We show that these proteins are required to form ILVs in Drosophila secondary cell recycling endosomes, but unlike core ESCRTs, they are not involved in degradation of ubiquitinylated proteins in late endosomes. Furthermore, CHMP5 knockdown in human HCT116 colorectal cancer cells selectively inhibits Rab11a-exosome production. Accessory ESCRT-III knockdown suppresses seminal fluid-mediated reproductive signalling by secondary cells and the growth-promoting activity of Rab11a-exosome-containing EVs from HCT116 cells. We conclude that accessory ESCRT-III components have a specific, ubiquitin-independent role in Rab11a-exosome generation, a mechanism that might be targeted to selectively block pro-tumorigenic activities of these vesicles in cancer.
Analysis in complementary fly and human models shows accessory ESCRT‐III components selectively regulate Rab11(a)‐exosome production in endosomes labelled by recycling endosomal marker Rab11(a). Furthermore, blocking this class of ESCRTs suppresses both pathological and physiological functions associated with exosomes.
Levels of accessory‐ESCRT‐III proteins are increased in small extracellular vesicle (sEV) preparations enriched with Rab11a‐exosomes; which are preferentially released following glutamine‐depletion of human cancer cell lines.
Accessory ESCRT‐III proteins are required for Rab11(a)‐exosome biogenesis in recycling endosomes, but not for ESCRT‐dependent processing of ubiquitinylated, late endosomal intraluminal vesicle cargos.
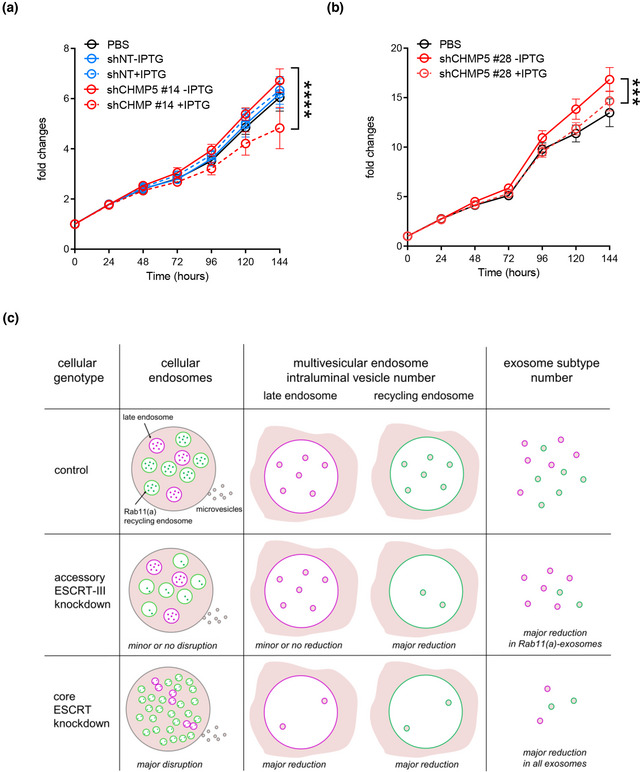
Accessory ESCRT‐III knockdown has a much milder effect on the cellular endolysosomal network than core ESCRT knockdown.
Knockdown of accessory ESCRT‐III proteins suppresses Rab11(a)‐exosome‐mediated human cancer cell growth and post‐mating responses in female flies.
Keywords: CHMP5; ESCRT; Rab11a-exosome; extracellular vesicle; recycling endosome.
© 2023 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Conflict of interest statement
The authors declare that they have no conflicts of interest in relation to this work.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
- 19076/CRUK_/Cancer Research UK/United Kingdom
- 097813/11/Z/WT_/Wellcome Trust/United Kingdom
- 107457/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- 091911/B/10/Z/WT_/Wellcome Trust/United Kingdom
- BB/R004862/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- C602/A18974/CRUK_/Cancer Research UK/United Kingdom
- BB/N016300/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/K017462/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- C19591/A19076/CRUK_/Cancer Research UK/United Kingdom
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
