

Genetic variation of morphological scaling in Drosophila melanogaster
- PMID: 36878946
- PMCID: PMC10162999
- DOI: 10.1038/s41437-023-00603-y
Genetic variation of morphological scaling in Drosophila melanogaster
Abstract
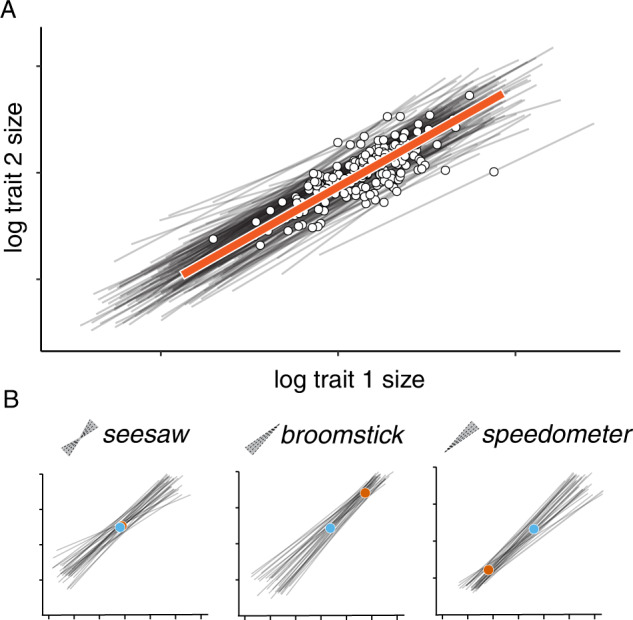
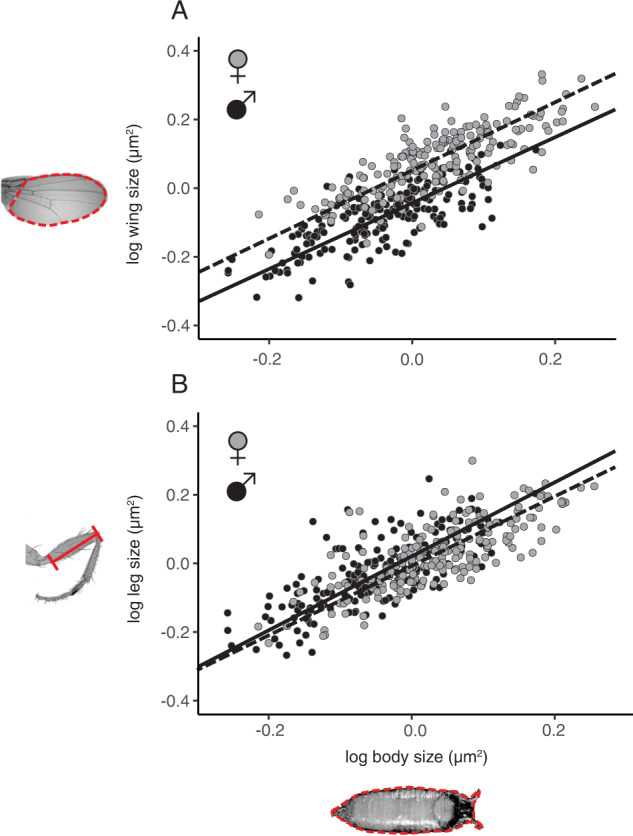
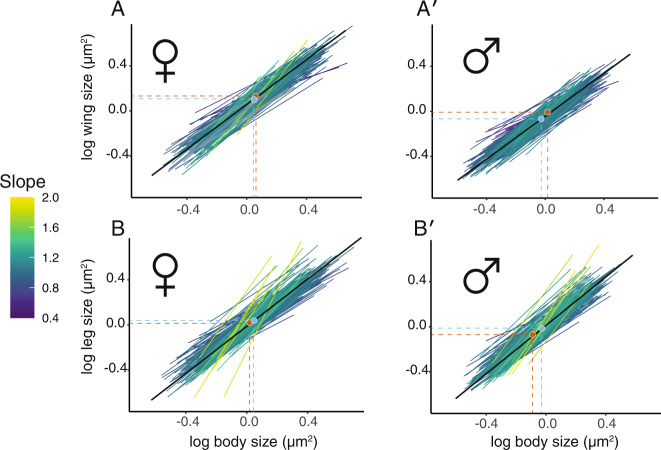
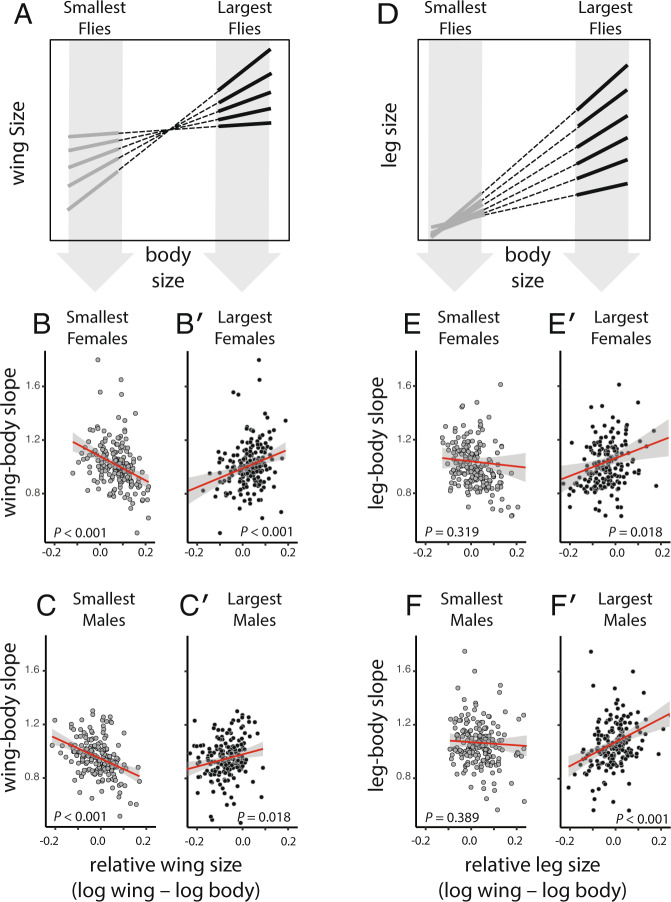
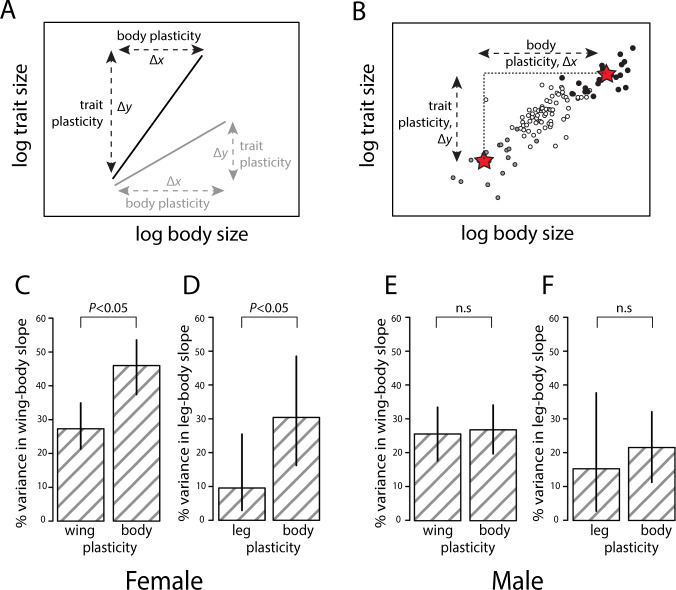
Morphological scaling relationships between the sizes of individual traits and the body captures the characteristic shape of a species, and their evolution is the primary mechanism of morphological diversification. However, we have almost no knowledge of the genetic variation of scaling, which is critical if we are to understand how scaling evolves. Here we explore the genetics of population scaling relationships (scaling relationships fit to multiple genetically-distinct individuals in a population) by describing the distribution of individual scaling relationships (genotype-specific scaling relationships that are unseen or cryptic). These individual scaling relationships harbor the genetic variation in the developmental mechanisms that regulate trait growth relative to body growth, and theoretical studies suggest that their distribution dictates how the population scaling relationship will respond to selection. Using variation in nutrition to generate size variation within 197 isogenic lineages of Drosophila melanogaster, we reveal extensive variation in the slopes of the wing-body and leg-body individual scaling relationships among genotypes. This variation reflects variation in the nutritionally-induced size plasticity of the wing, leg, and body. Surprisingly, we find that variation in the slope of individual scaling relationships primarily results from variation in nutritionally-induced plasticity of body size, not leg or wing size. These data allow us to predict how different selection regimes affect scaling in Drosophila, and is the first step in identifying the genetic targets of such selection. More generally, our approach provides a framework for understanding the genetic variation of scaling, an important prerequisite to explaining how selection changes scaling and morphology.
© 2023. The Author(s), under exclusive licence to The Genetics Society.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Baker RH, Wilkinson GS. Phylogenetic analysis of sexual dimorphism and eye-span allometry in stalk-eyed flies (Diopsidae) Evolution. 2001;55:1373–1385. - PubMed
-
- Bates D, Mächler M, Bolker B, Walker S (2015). Fitting Linear Mixed-Effects Models Using lme4. J Stat Softw 67
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
