

Medicinal plant-derived mtDNA via nanovesicles induces the cGAS-STING pathway to remold tumor-associated macrophages for tumor regression
- PMID: 36879291
- PMCID: PMC9990354
- DOI: 10.1186/s12951-023-01835-0
Medicinal plant-derived mtDNA via nanovesicles induces the cGAS-STING pathway to remold tumor-associated macrophages for tumor regression
Abstract
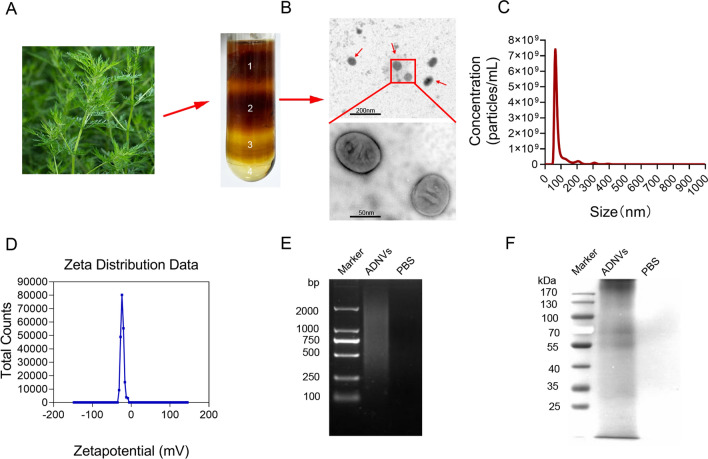
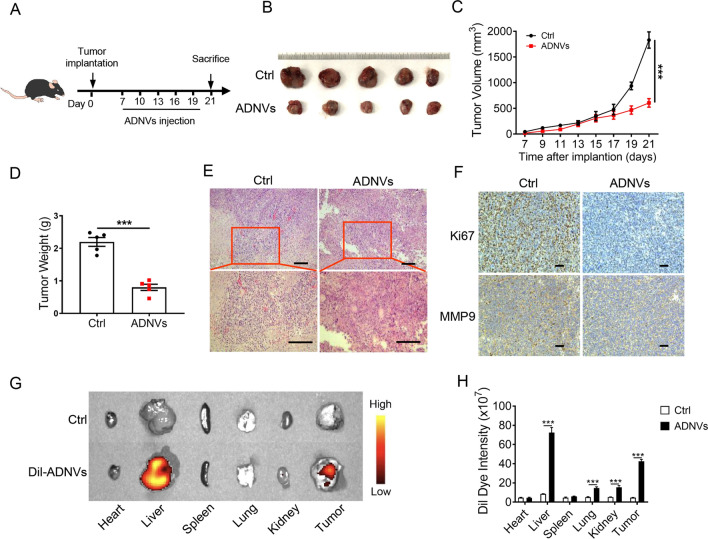
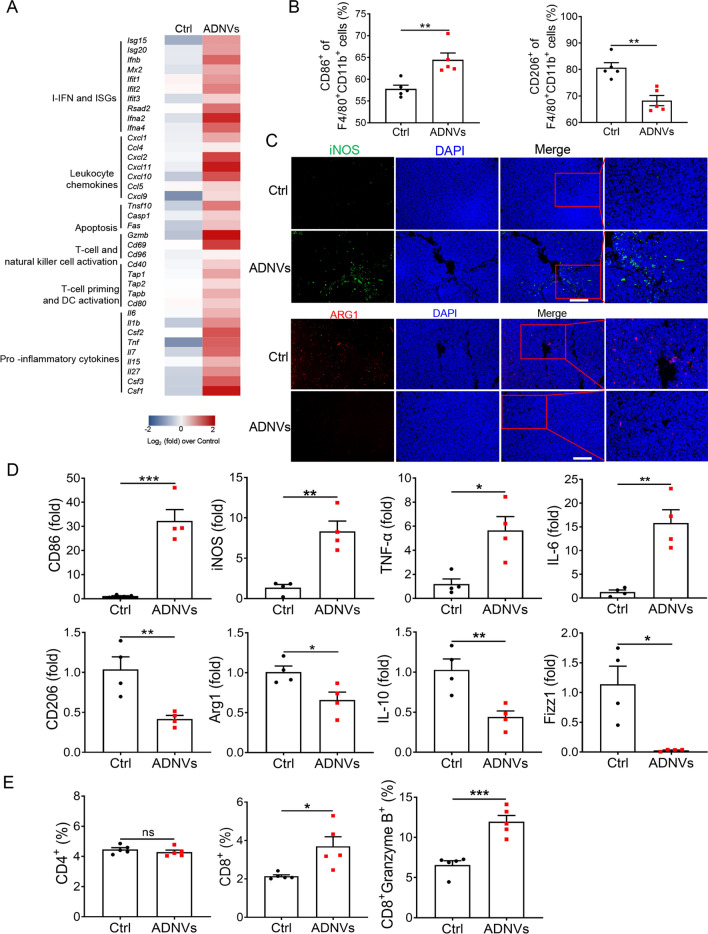
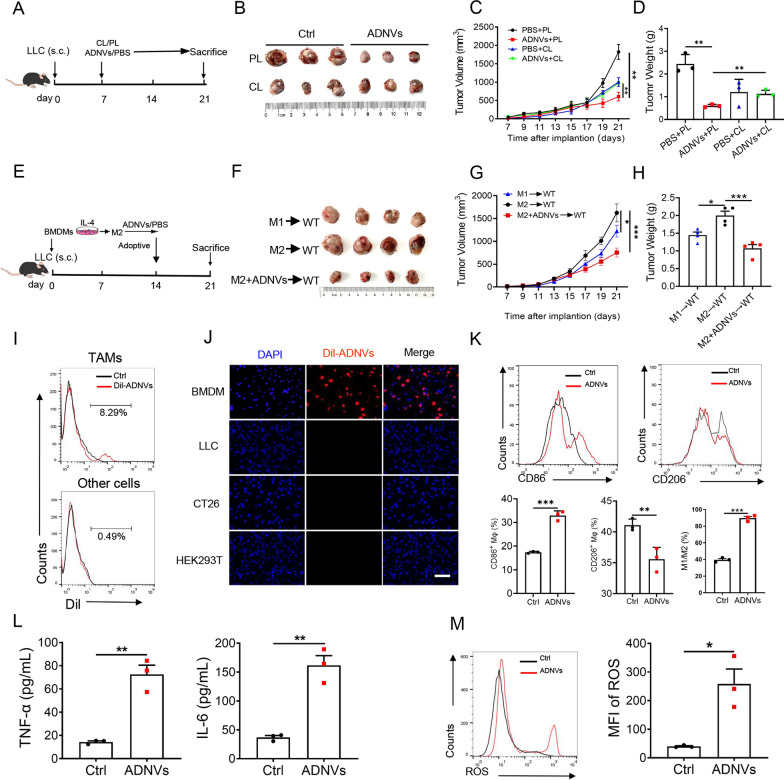
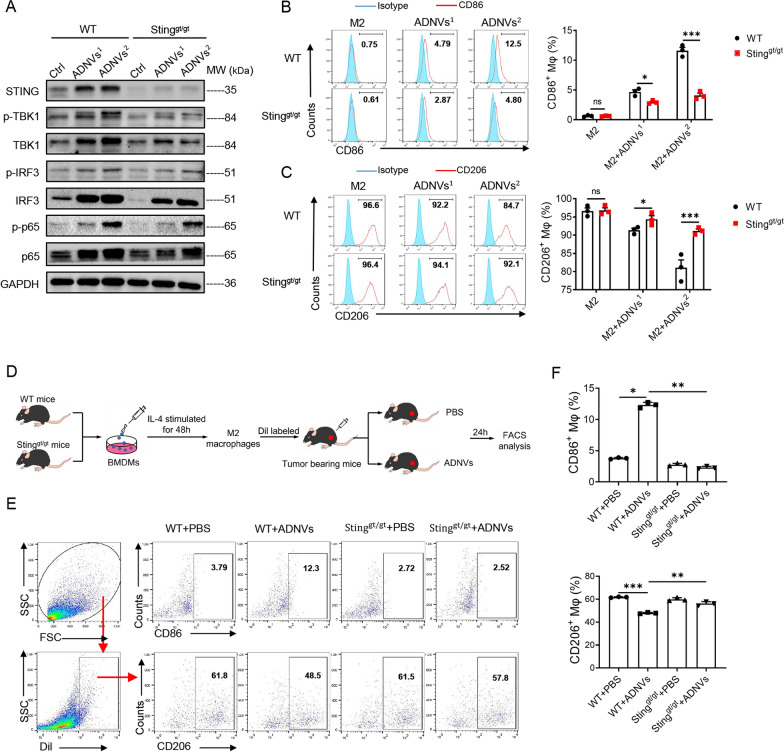
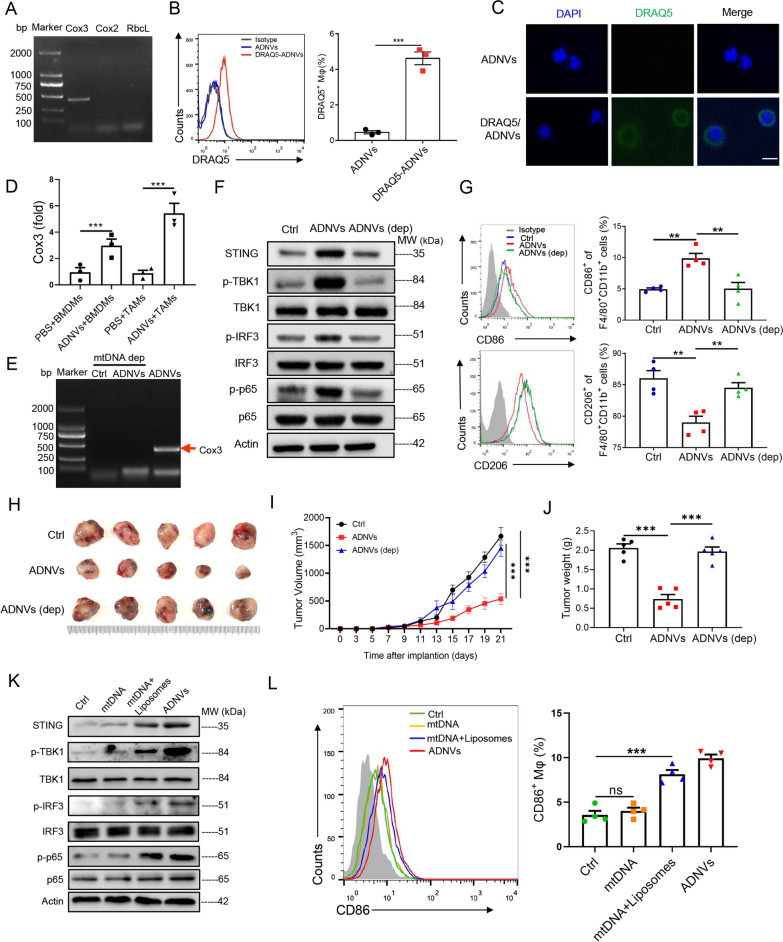
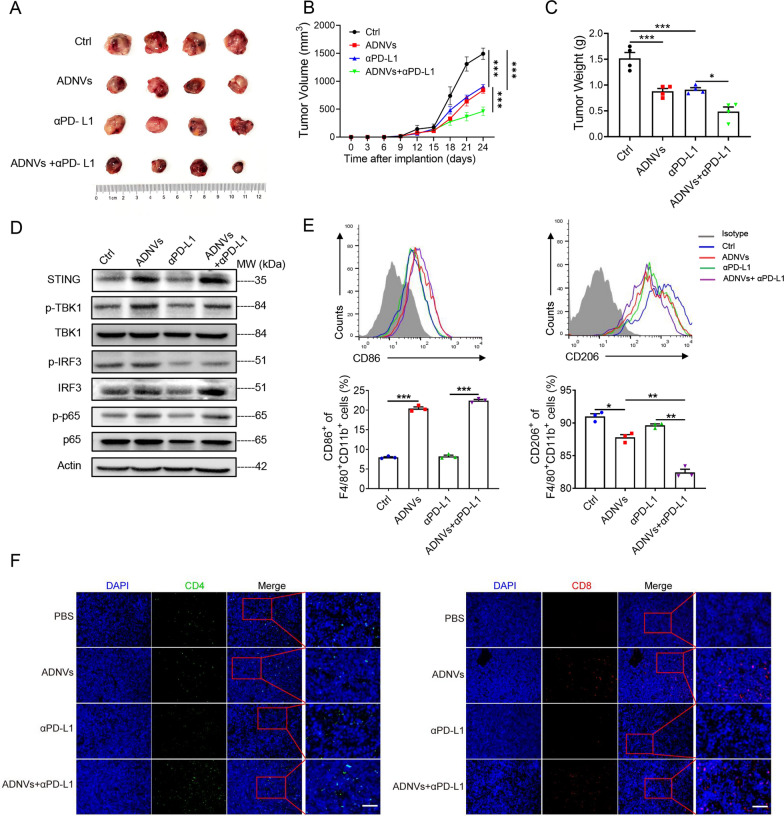
Plant-derived nanovesicles (PDNVs) have been proposed as a major mechanism for the inter-kingdom interaction and communication, but the effector components enclosed in the vesicles and the mechanisms involved are largely unknown. The plant Artemisia annua is known as an anti-malaria agent that also exhibits a wide range of biological activities including the immunoregulatory and anti-tumor properties with the mechanisms to be further addressed. Here, we isolated and purified the exosome-like particles from A. annua, which were characterized by nano-scaled and membrane-bound shape and hence termed artemisia-derived nanovesicles (ADNVs). Remarkably, the vesicles demonstrated to inhibit tumor growth and boost anti-tumor immunity in a mouse model of lung cancer, primarily through remolding the tumor microenvironment and reprogramming tumor-associated macrophages (TAMs). We identified plant-derived mitochondrial DNA (mtDNA), upon internalized into TAMs via the vesicles, as a major effector molecule to induce the cGAS-STING pathway driving the shift of pro-tumor macrophages to anti-tumor phenotype. Furthermore, our data showed that administration of ADNVs greatly improved the efficacy of PD-L1 inhibitor, a prototypic immune checkpoint inhibitor, in tumor-bearing mice. Together, the present study, for the first time, to our knowledge, unravels an inter-kingdom interaction wherein the medical plant-derived mtDNA, via the nanovesicles, induces the immunostimulatory signaling in mammalian immune cells for resetting anti-tumor immunity and promoting tumor eradication.
Keywords: Artemisia-derived nanovesicles; Tumor-associated macrophages; cGAS-STING; mtDNA.
© 2023. The Author(s).
Conflict of interest statement
The authors have no competing of interest to declare.
Figures
Similar articles
-
Modulation of alveolar macrophage and mitochondrial fitness by medicinal plant-derived nanovesicles to mitigate acute lung injury and viral pneumonia.J Nanobiotechnology. 2024 Apr 18;22(1):190. doi: 10.1186/s12951-024-02473-w. J Nanobiotechnology. 2024. PMID: 38637808 Free PMC article.
-
Mitochondrial DNA leakage induces odontoblast inflammation via the cGAS-STING pathway.Cell Commun Signal. 2021 May 20;19(1):58. doi: 10.1186/s12964-021-00738-7. Cell Commun Signal. 2021. PMID: 34016129 Free PMC article.
-
Salmonella Induces the cGAS-STING-Dependent Type I Interferon Response in Murine Macrophages by Triggering mtDNA Release.mBio. 2022 Jun 28;13(3):e0363221. doi: 10.1128/mbio.03632-21. Epub 2022 May 23. mBio. 2022. PMID: 35604097 Free PMC article.
-
cGAS/cGAMP/STING signal propagation in the tumor microenvironment: Key role for myeloid cells in antitumor immunity.Radiother Oncol. 2022 Sep;174:158-167. doi: 10.1016/j.radonc.2022.07.014. Epub 2022 Jul 20. Radiother Oncol. 2022. PMID: 35870728 Review.
-
Herbal Medicine-Derived Exosome-Like Nanovesicles: A Rising Star in Cancer Therapy.Int J Nanomedicine. 2024 Jul 25;19:7585-7603. doi: 10.2147/IJN.S477270. eCollection 2024. Int J Nanomedicine. 2024. PMID: 39081899 Free PMC article. Review.
Cited by
-
Nano-strategies for Targeting Tumor-Associated Macrophages in Cancer immunotherapy.J Cancer. 2025 Mar 31;16(7):2261-2274. doi: 10.7150/jca.108194. eCollection 2025. J Cancer. 2025. PMID: 40302816 Free PMC article. Review.
-
Mechanistic Insights into the Anti-Glioma Effects of Exosome-Like Nanoparticles Derived from Garcinia Mangostana L.: A Metabolomics, Network Pharmacology, and Experimental Study.Int J Nanomedicine. 2025 Apr 27;20:5407-5427. doi: 10.2147/IJN.S514930. eCollection 2025. Int J Nanomedicine. 2025. PMID: 40321800 Free PMC article.
-
Harnessing innate immune pathways for therapeutic advancement in cancer.Signal Transduct Target Ther. 2024 Mar 25;9(1):68. doi: 10.1038/s41392-024-01765-9. Signal Transduct Target Ther. 2024. PMID: 38523155 Free PMC article. Review.
-
Mutual regulation of PD-L1 immunosuppression between tumor-associated macrophages and tumor cells: a critical role for exosomes.Cell Commun Signal. 2024 Jan 9;22(1):21. doi: 10.1186/s12964-024-01473-5. Cell Commun Signal. 2024. PMID: 38195554 Free PMC article. Review.
-
Plant-derived nanovesicles: harnessing nature's power for tissue protection and repair.J Nanobiotechnology. 2023 Nov 24;21(1):445. doi: 10.1186/s12951-023-02193-7. J Nanobiotechnology. 2023. PMID: 38001440 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
