

Vitamin A regulates tissue-specific organ remodeling in diet-induced obesity independent of mitochondrial function
- PMID: 36891060
- PMCID: PMC9987331
- DOI: 10.3389/fendo.2023.1118751
Vitamin A regulates tissue-specific organ remodeling in diet-induced obesity independent of mitochondrial function
Abstract
Background: Perturbed mitochondrial energetics and vitamin A (VitA) metabolism are associated with the pathogenesis of diet-induced obesity (DIO) and type 2 diabetes (T2D).
Methods: To test the hypothesis that VitA regulates tissue-specific mitochondrial energetics and adverse organ remodeling in DIO, we utilized a murine model of impaired VitA availability and high fat diet (HFD) feeding. Mitochondrial respiratory capacity and organ remodeling were assessed in liver, skeletal muscle, and kidney tissue, which are organs affected by T2D-associated complications and are critical for the pathogenesis of T2D.
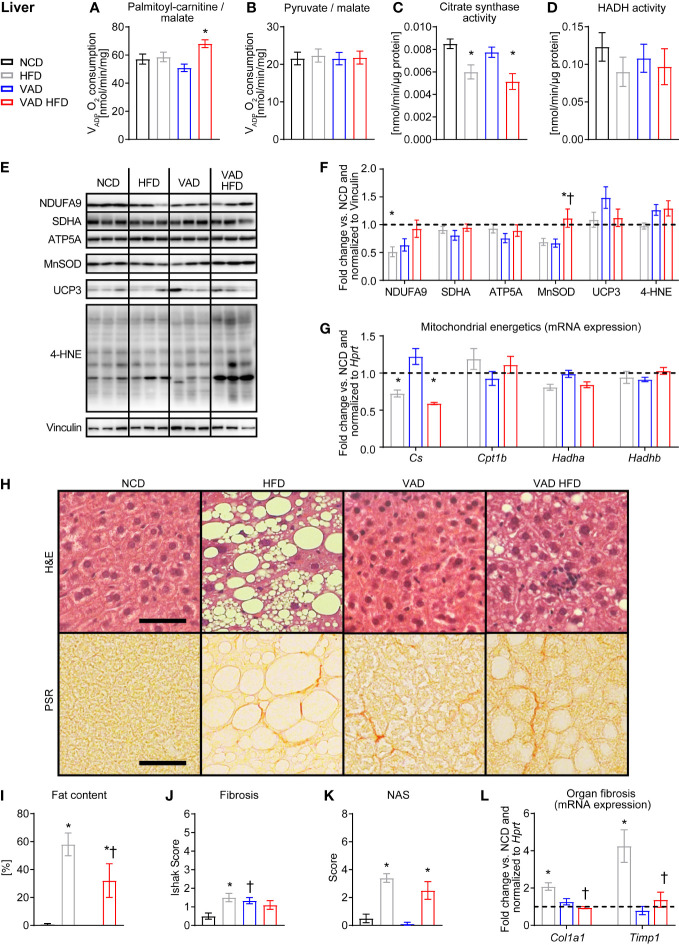
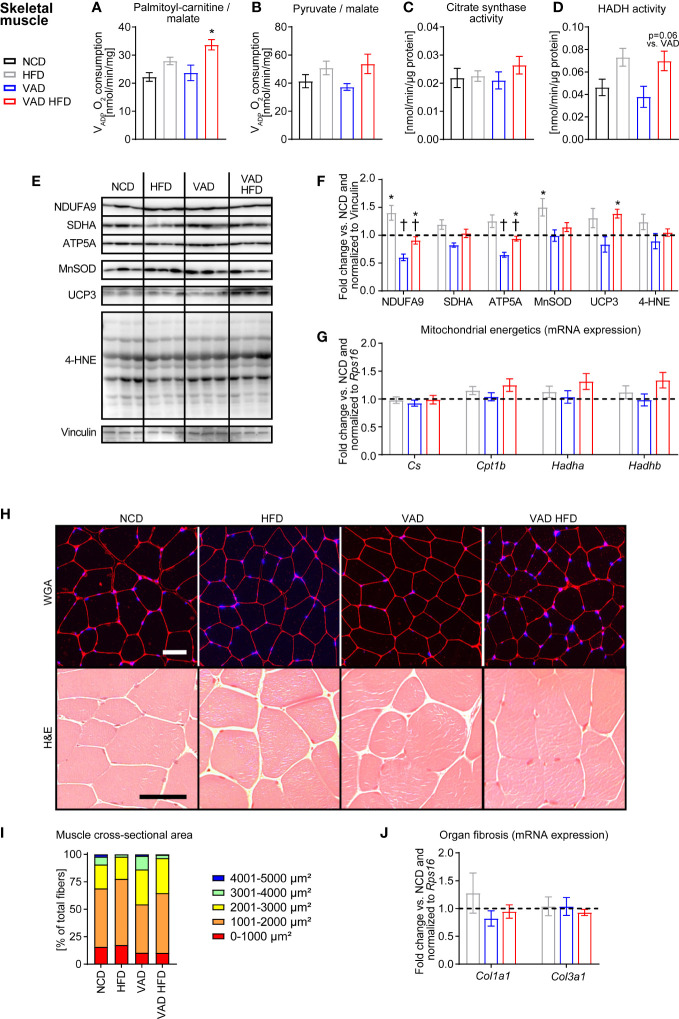
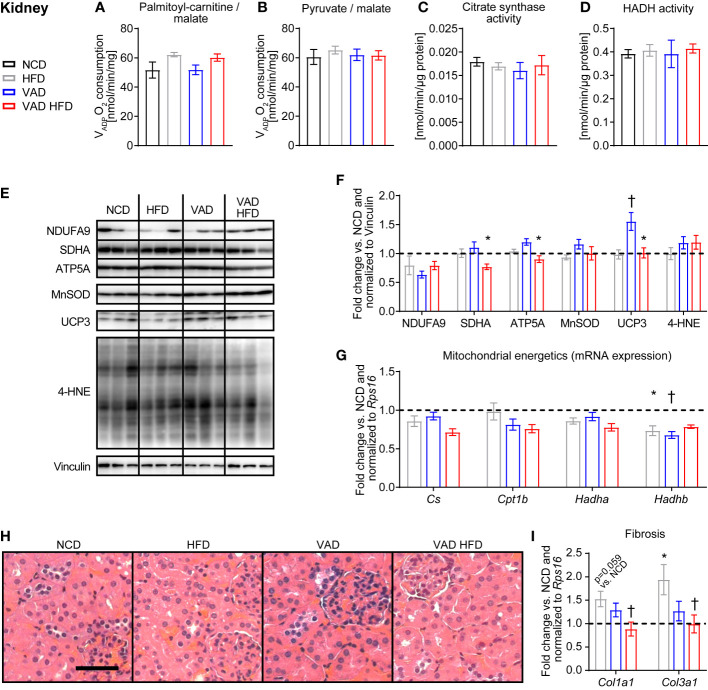
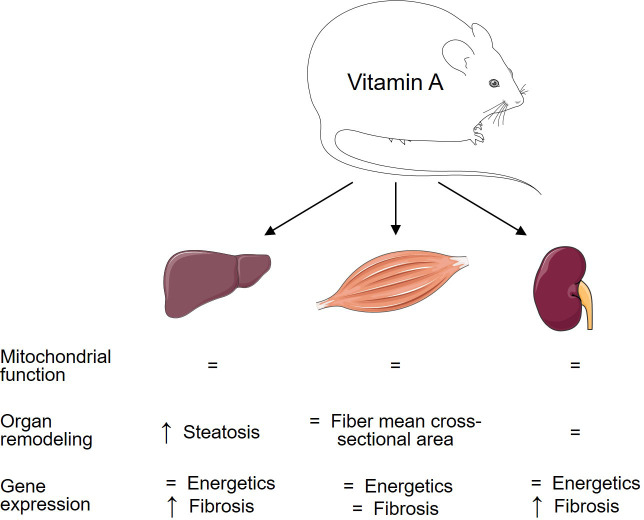
Results: In liver, VitA had no impact on maximal ADP-stimulated mitochondrial respiratory capacity (VADP) following HFD feeding with palmitoyl-carnitine and pyruvate each combined with malate as substrates. Interestingly, histopathological and gene expression analyses revealed that VitA mediates steatosis and adverse remodeling in DIO. In skeletal muscle, VitA did not affect VADP following HFD feeding. No morphological differences were detected between groups. In kidney, VADP was not different between groups with both combinations of substrates and VitA transduced the pro-fibrotic transcriptional response following HFD feeding.
Conclusion: The present study identifies an unexpected and tissue-specific role for VitA in DIO that regulates the pro-fibrotic transcriptional response and that results in organ damage independent of changes in mitochondrial energetics.
Keywords: diet-induced obesity; kidney; liver; mitochondria; skeletal muscle; type 2 diabetes; vitamin A.
Copyright © 2023 Shymotiuk, Froese, Werlein, Naasner, Szaroszyk, Kühnel, Jonigk, Blaner, Wende, Abel, Bauersachs and Riehle.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases