

The connectome of an insect brain
- PMID: 36893230
- PMCID: PMC7614541
- DOI: 10.1126/science.add9330
The connectome of an insect brain
Abstract
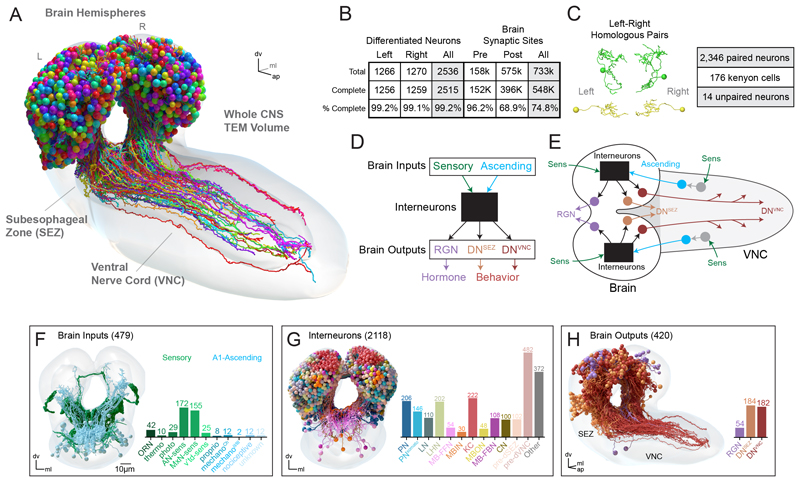
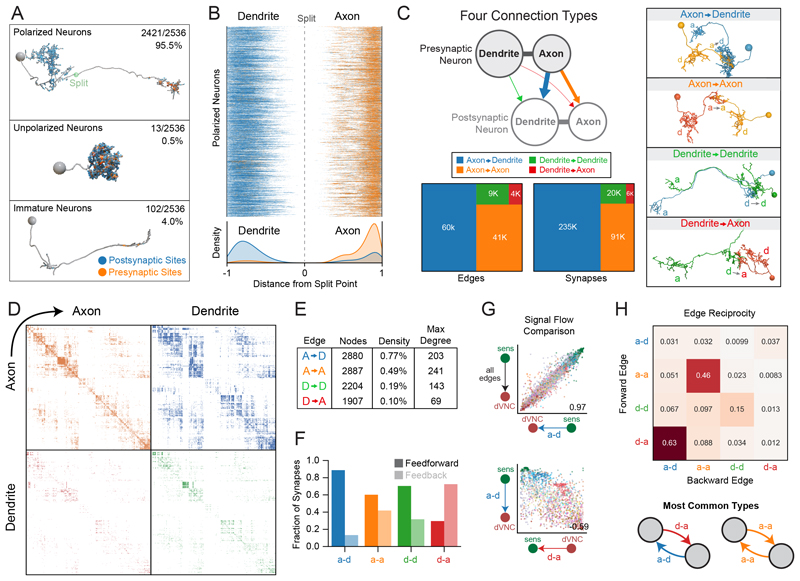
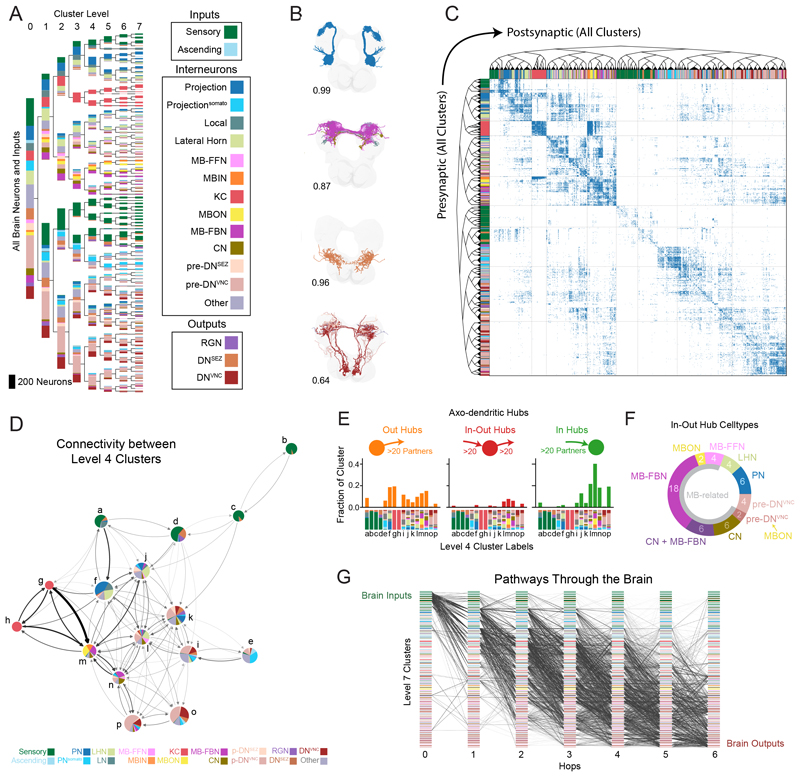
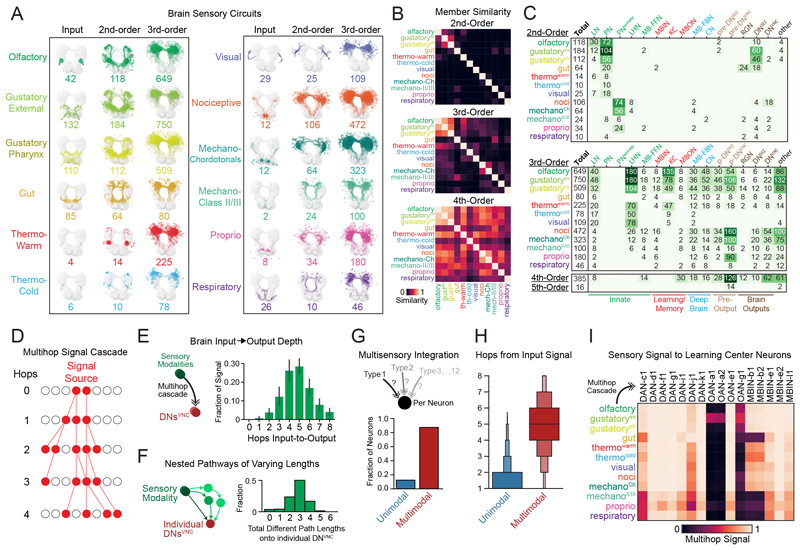
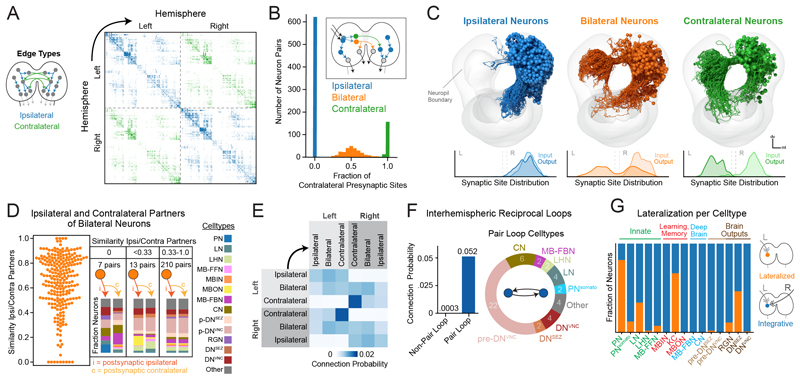
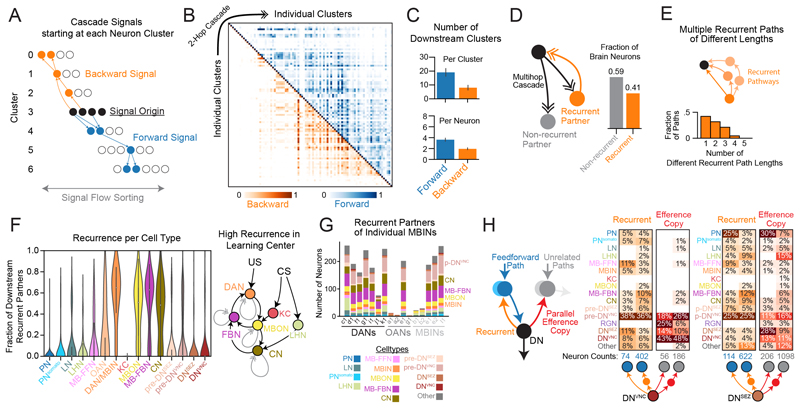
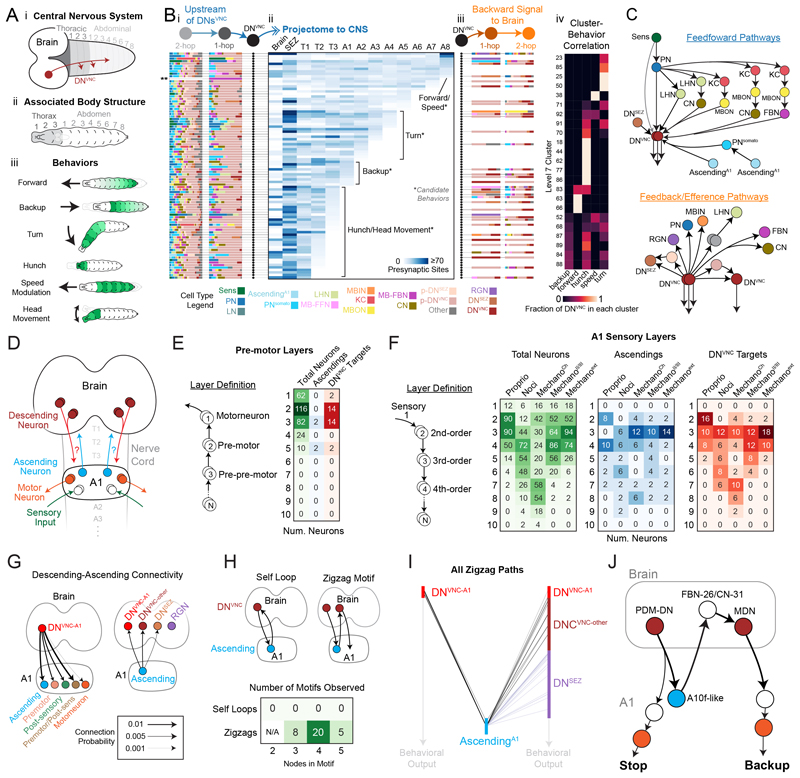
Brains contain networks of interconnected neurons and so knowing the network architecture is essential for understanding brain function. We therefore mapped the synaptic-resolution connectome of an entire insect brain (Drosophila larva) with rich behavior, including learning, value computation, and action selection, comprising 3016 neurons and 548,000 synapses. We characterized neuron types, hubs, feedforward and feedback pathways, as well as cross-hemisphere and brain-nerve cord interactions. We found pervasive multisensory and interhemispheric integration, highly recurrent architecture, abundant feedback from descending neurons, and multiple novel circuit motifs. The brain's most recurrent circuits comprised the input and output neurons of the learning center. Some structural features, including multilayer shortcuts and nested recurrent loops, resembled state-of-the-art deep learning architectures. The identified brain architecture provides a basis for future experimental and theoretical studies of neural circuits.
Conflict of interest statement
Figures
Comment in
-
Gigantic map of fly brain is a first for a complex animal.Nature. 2023 Mar;615(7953):571. doi: 10.1038/d41586-023-00709-7. Nature. 2023. PMID: 36899190 No abstract available.
References
-
- Verasztó C, et al. bioRxiv. 2020:2020.08.21.260984. doi: 10.1101/2020.08.21.260984. Preprint. - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
