

Divalent siRNAs are bioavailable in the lung and efficiently block SARS-CoV-2 infection
- PMID: 36893269
- PMCID: PMC10089225
- DOI: 10.1073/pnas.2219523120
Divalent siRNAs are bioavailable in the lung and efficiently block SARS-CoV-2 infection
Abstract
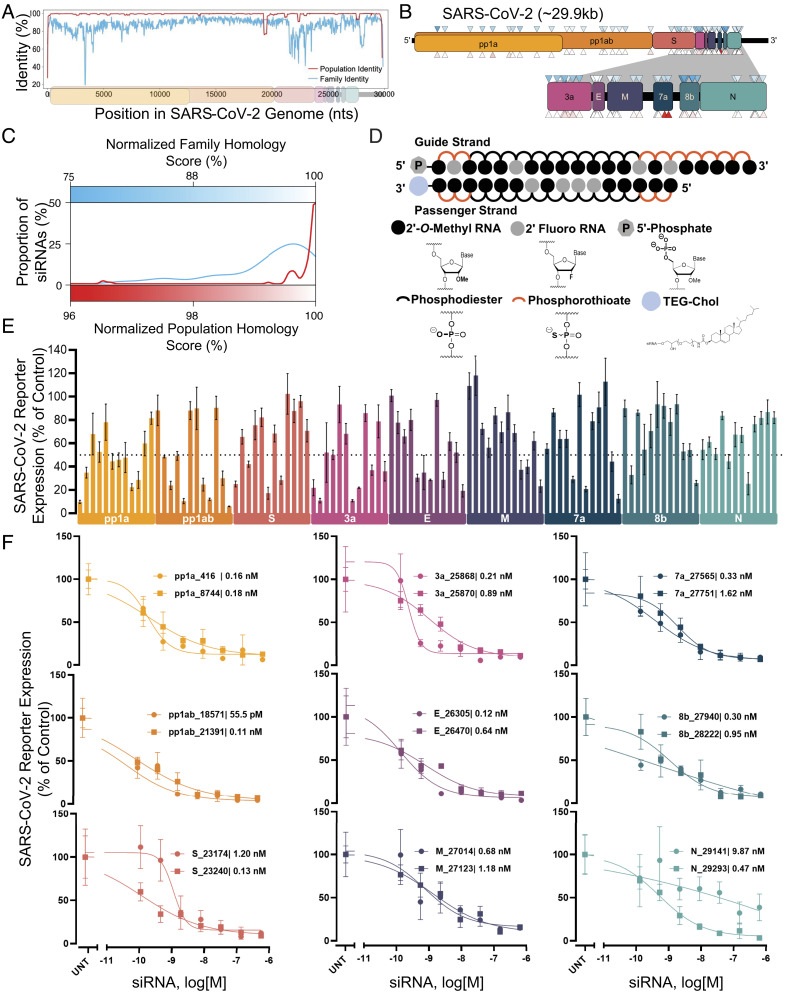
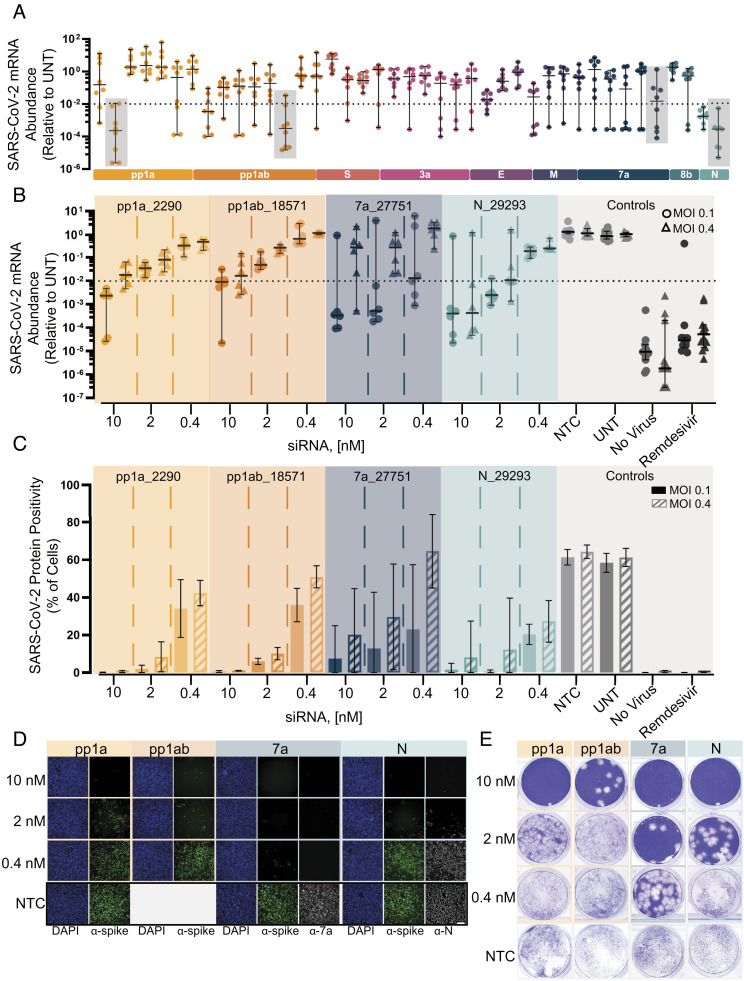
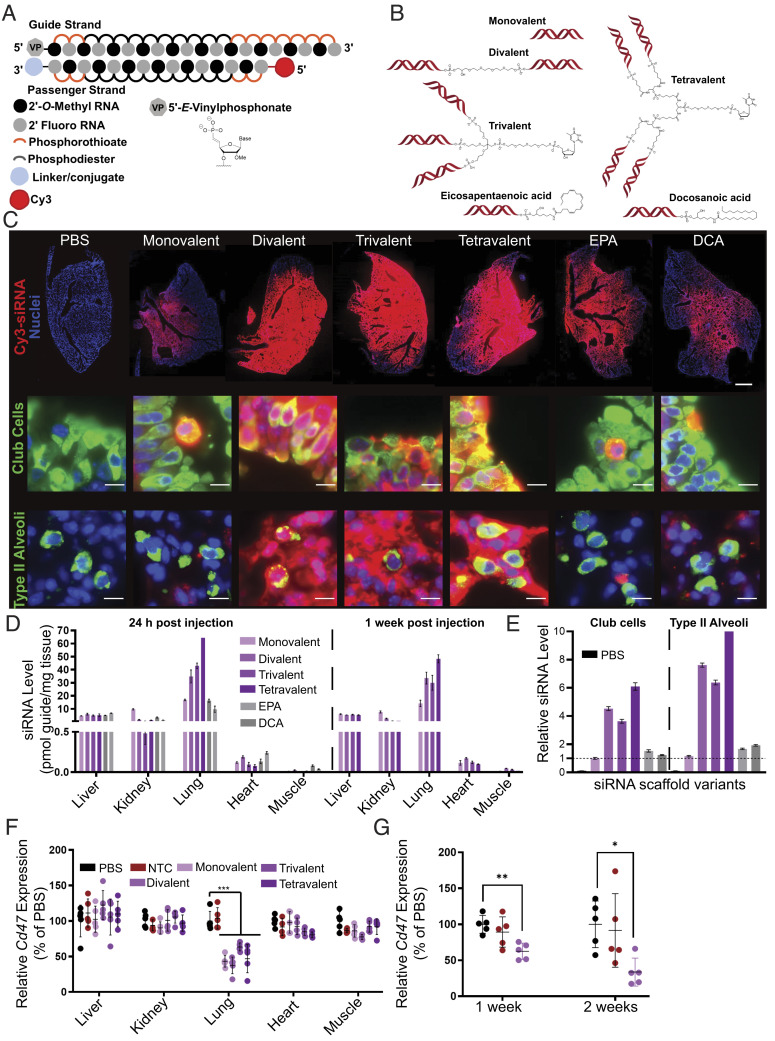
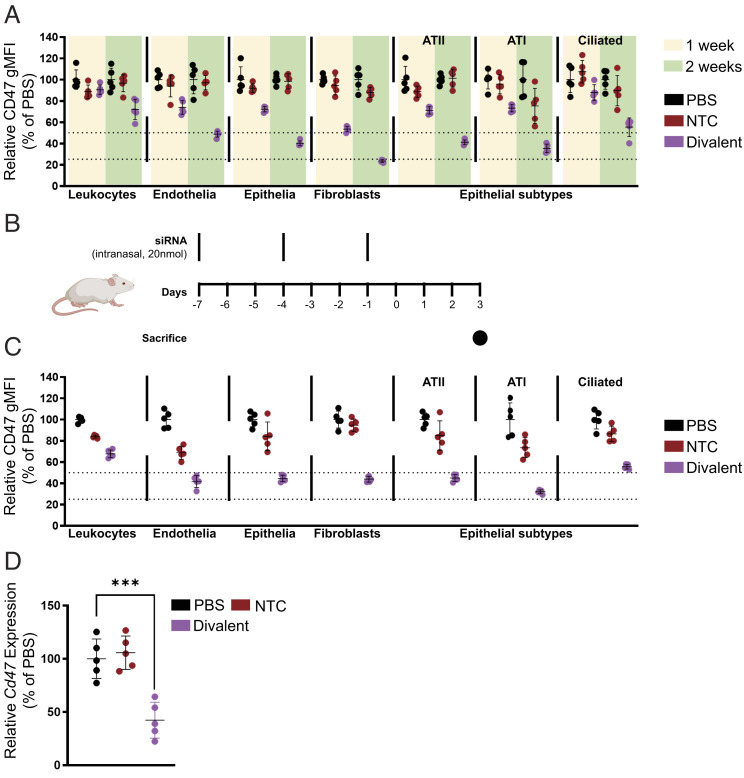
The continuous evolution of SARS-CoV-2 variants complicates efforts to combat the ongoing pandemic, underscoring the need for a dynamic platform for the rapid development of pan-viral variant therapeutics. Oligonucleotide therapeutics are enhancing the treatment of numerous diseases with unprecedented potency, duration of effect, and safety. Through the systematic screening of hundreds of oligonucleotide sequences, we identified fully chemically stabilized siRNAs and ASOs that target regions of the SARS-CoV-2 genome conserved in all variants of concern, including delta and omicron. We successively evaluated candidates in cellular reporter assays, followed by viral inhibition in cell culture, with eventual testing of leads for in vivo antiviral activity in the lung. Previous attempts to deliver therapeutic oligonucleotides to the lung have met with only modest success. Here, we report the development of a platform for identifying and generating potent, chemically modified multimeric siRNAs bioavailable in the lung after local intranasal and intratracheal delivery. The optimized divalent siRNAs showed robust antiviral activity in human cells and mouse models of SARS-CoV-2 infection and represent a new paradigm for antiviral therapeutic development for current and future pandemics.
Keywords: SARS-COV-2; antisense oligos; lung; mouse model; small interfering RNAs.
Conflict of interest statement
The authors have filed a patent application related to this work. G. Harold and Leila Y. Mathers Foundation (Mathers Foundation), UH | Harrington Discovery Institute, University Hospitals (HDI), HHS | NIH, HHS | NIH, HHS | NIH, Larry Ellison Foundation (LEF), Ono Pharmaceutical Foundation, Massachusetts Consortium on Pathogen Readiness (MCPR), U.S. Department of Defense (DOD), University of Massachusetts Chan Medical School, University of Massachusetts Chan Medical School.
Figures
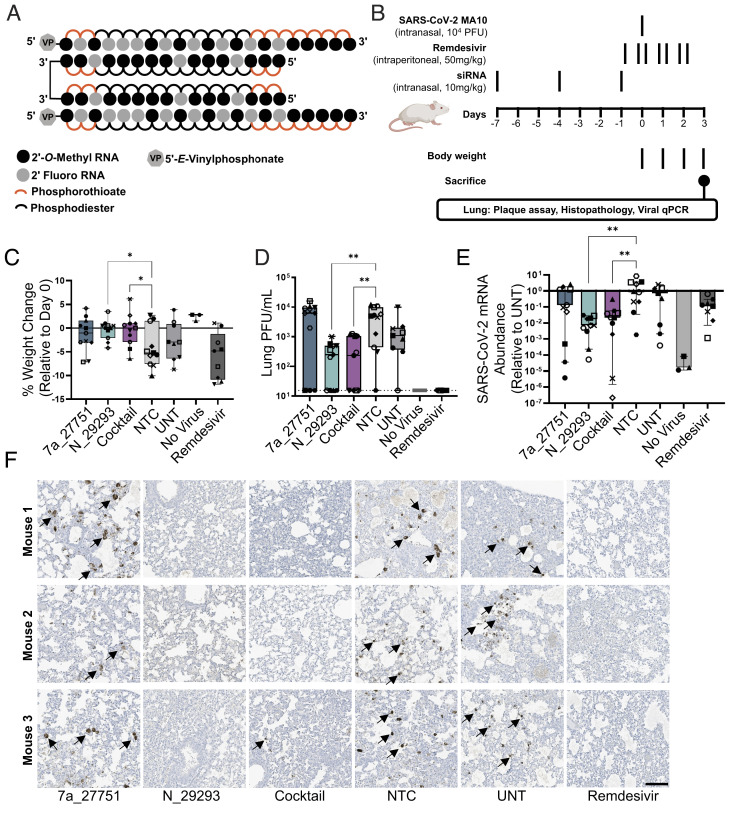
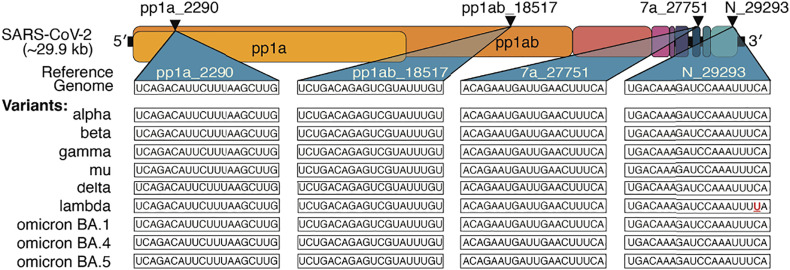
References
-
- Chakraborty C., Bhattacharya M., Sharma A. R., Present variants of concern and variants of interest of severe acute respiratory syndrome coronavirus 2: Their significant mutations in S-glycoprotein, infectivity, re-infectivity, immune escape and vaccines activity. Rev. Med. Virol. 32, e2270.
-
- Lauring A. S., Hodcroft E. B., Genetic variants of SARS-CoV-2-what do they mean? JAMA 325, 529–531 (2021). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
