

Broader functions of TIR domains in Arabidopsis immunity
- PMID: 36893276
- PMCID: PMC10242710
- DOI: 10.1073/pnas.2220921120
Broader functions of TIR domains in Arabidopsis immunity
Abstract
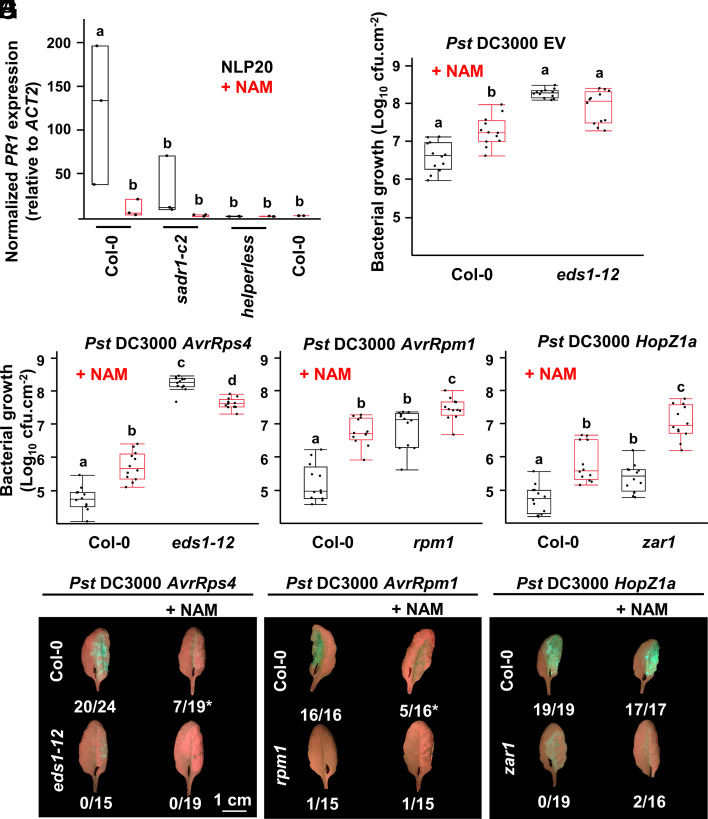
TIR domains are NAD-degrading enzymes that function during immune signaling in prokaryotes, plants, and animals. In plants, most TIR domains are incorporated into intracellular immune receptors termed TNLs. In Arabidopsis, TIR-derived small molecules bind and activate EDS1 heterodimers, which in turn activate RNLs, a class of cation channel-forming immune receptors. RNL activation drives cytoplasmic Ca2+ influx, transcriptional reprogramming, pathogen resistance, and host cell death. We screened for mutants that suppress an RNL activation mimic allele and identified a TNL, SADR1. Despite being required for the function of an autoactivated RNL, SADR1 is not required for defense signaling triggered by other tested TNLs. SADR1 is required for defense signaling initiated by some transmembrane pattern recognition receptors and contributes to the unbridled spread of cell death in lesion simulating disease 1. Together with RNLs, SADR1 regulates defense gene expression at infection site borders, likely in a non-cell autonomous manner. RNL mutants that cannot sustain this pattern of gene expression are unable to prevent disease spread beyond localized infection sites, suggesting that this pattern corresponds to a pathogen containment mechanism. SADR1 potentiates RNL-driven immune signaling not only through the activation of EDS1 but also partially independently of EDS1. We studied EDS1-independent TIR function using nicotinamide, an NADase inhibitor. Nicotinamide decreased defense induction from transmembrane pattern recognition receptors and decreased calcium influx, pathogen growth restriction, and host cell death following intracellular immune receptor activation. We demonstrate that TIR domains can potentiate calcium influx and defense and are thus broadly required for Arabidopsis immunity.
Keywords: Arabidopsis; NLR; TIR domains; immunity.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
-
- Essuman K., Milbrandt J., Dangl J. L., Nishimura M. T., Shared TIR enzymatic functions regulate cell death and immunity across the tree of life. Science 377, eabo0001 (2022). - PubMed
-
- Yu D., et al. , TIR domains of plant immune receptors are 2’,3’-cAMP/cGMP synthetases mediating cell death. Cell 185, 2370–2386.e18 (2022). - PubMed
-
- Jia A., et al. , TIR-catalyzed ADP-ribosylation reactions produce signaling molecules for plant immunity. Science 377 (2022). - PubMed
-
- Huang S., et al. , Identification and receptor mechanism of TIR-catalyzed small molecules in plant immunity. Science 377, eabq3297 (2022). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
