

Nanopore Detection Using Supercharged Polypeptide Molecular Carriers
- PMID: 36897933
- PMCID: PMC10037339
- DOI: 10.1021/jacs.2c13465
Nanopore Detection Using Supercharged Polypeptide Molecular Carriers
Abstract
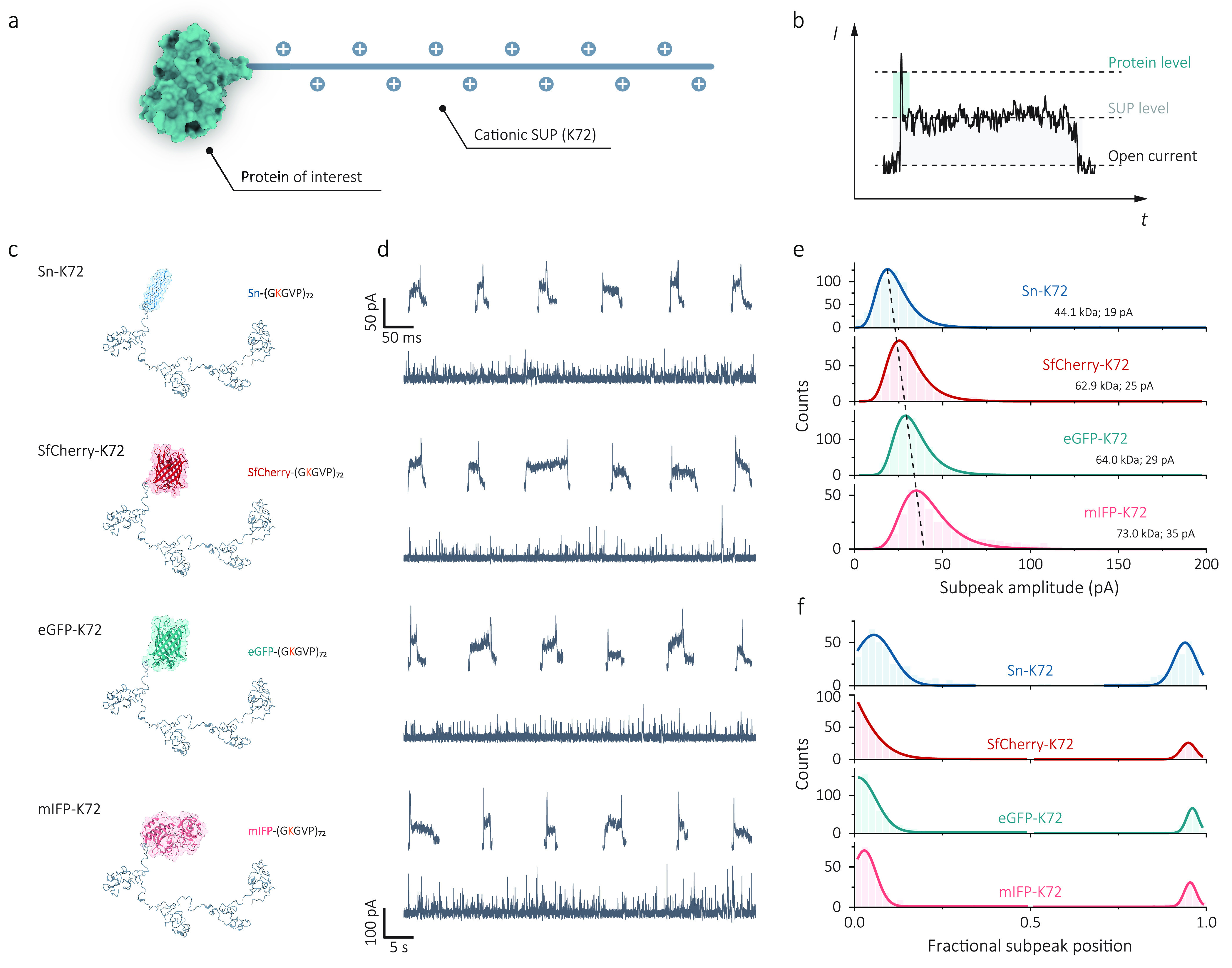
The analysis at the single-molecule level of proteins and their interactions can provide critical information for understanding biological processes and diseases, particularly for proteins present in biological samples with low copy numbers. Nanopore sensing is an analytical technique that allows label-free detection of single proteins in solution and is ideally suited to applications, such as studying protein-protein interactions, biomarker screening, drug discovery, and even protein sequencing. However, given the current spatiotemporal limitations in protein nanopore sensing, challenges remain in controlling protein translocation through a nanopore and relating protein structures and functions with nanopore readouts. Here, we demonstrate that supercharged unstructured polypeptides (SUPs) can be genetically fused with proteins of interest and used as molecular carriers to facilitate nanopore detection of proteins. We show that cationic SUPs can substantially slow down the translocation of target proteins due to their electrostatic interactions with the nanopore surface. This approach enables the differentiation of individual proteins with different sizes and shapes via characteristic subpeaks in the nanopore current, thus facilitating a viable route to use polypeptide molecular carriers to control molecular transport and as a potential system to study protein-protein interactions at the single-molecule level.
Conflict of interest statement
The authors declare no competing financial interest.
Figures




References
-
- Xue L.; Yamazaki H.; Ren R.; Wanunu M.; Ivanov A. P.; Edel J. B. Solid-State Nanopore Sensors. Nat. Rev. Mater. 2020, 5, 931.10.1038/s41578-020-0229-6. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
