

TNIP1 inhibits selective autophagy via bipartite interaction with LC3/GABARAP and TAX1BP1
- PMID: 36898370
- PMCID: PMC10112281
- DOI: 10.1016/j.molcel.2023.02.023
TNIP1 inhibits selective autophagy via bipartite interaction with LC3/GABARAP and TAX1BP1
Abstract
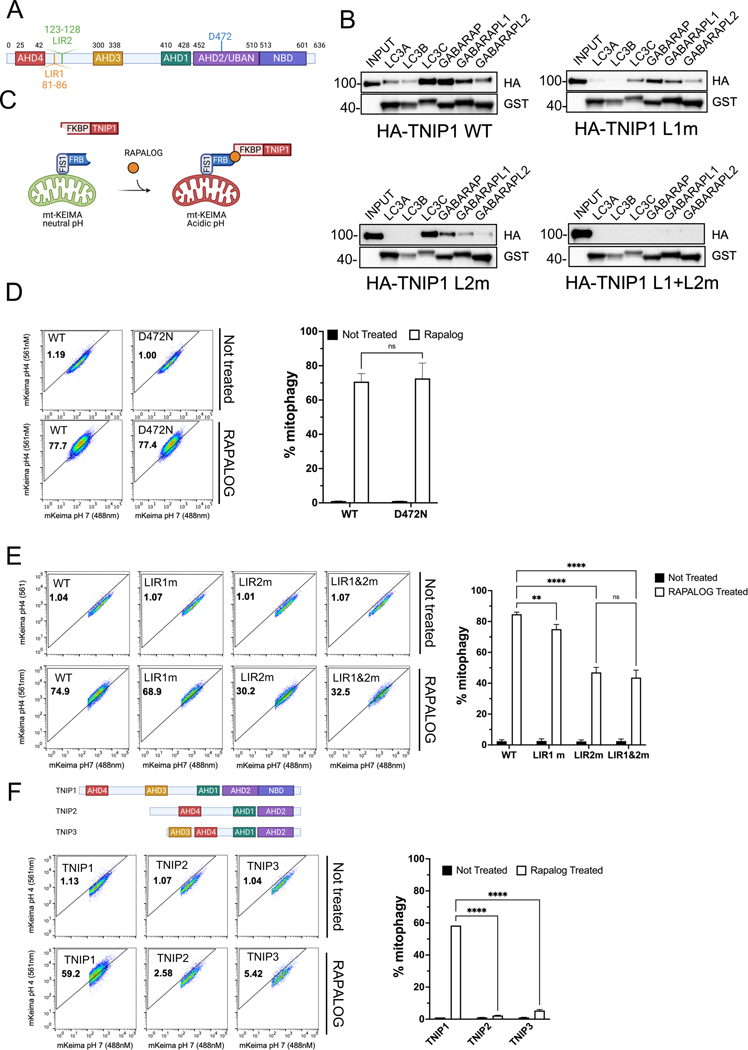
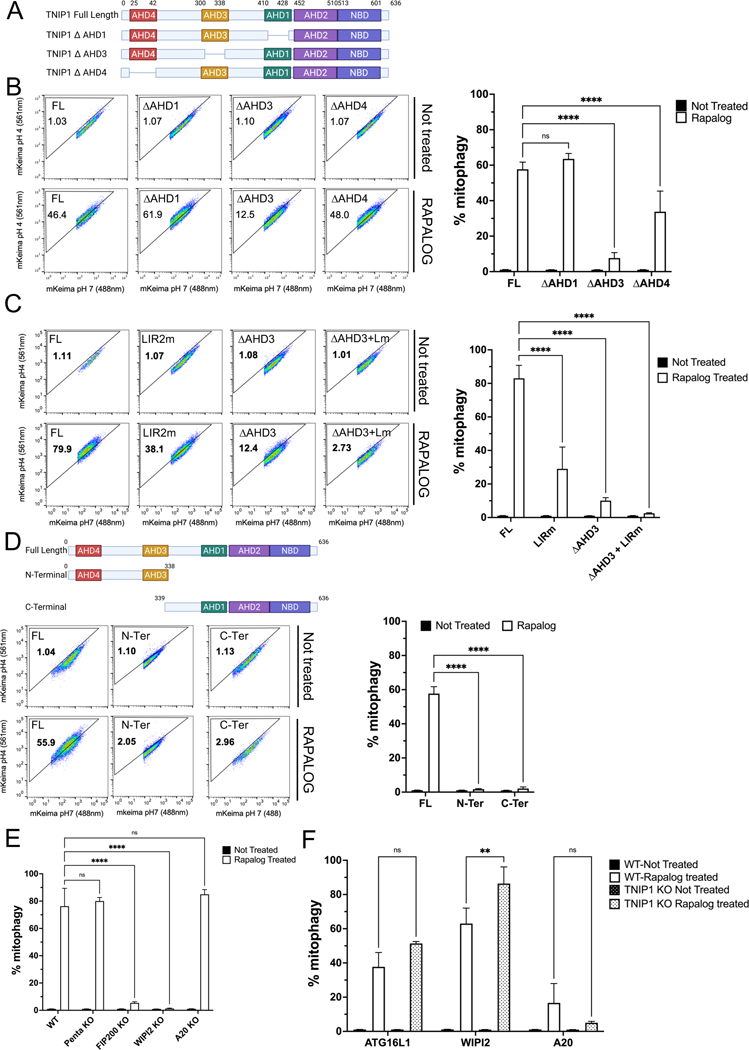
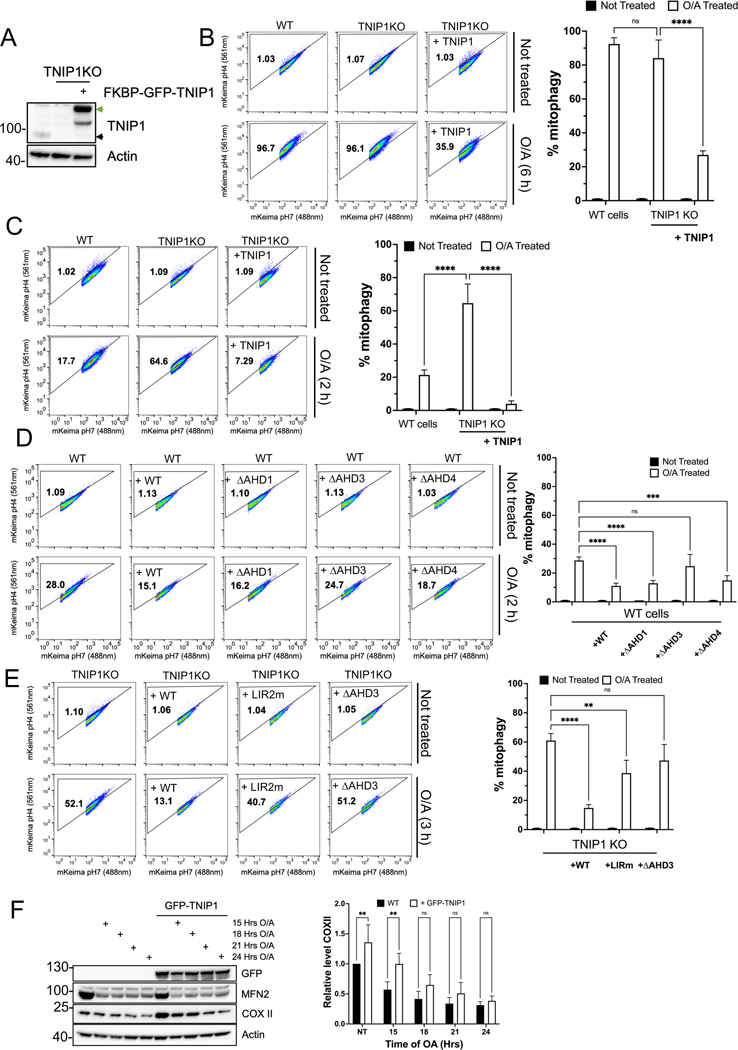
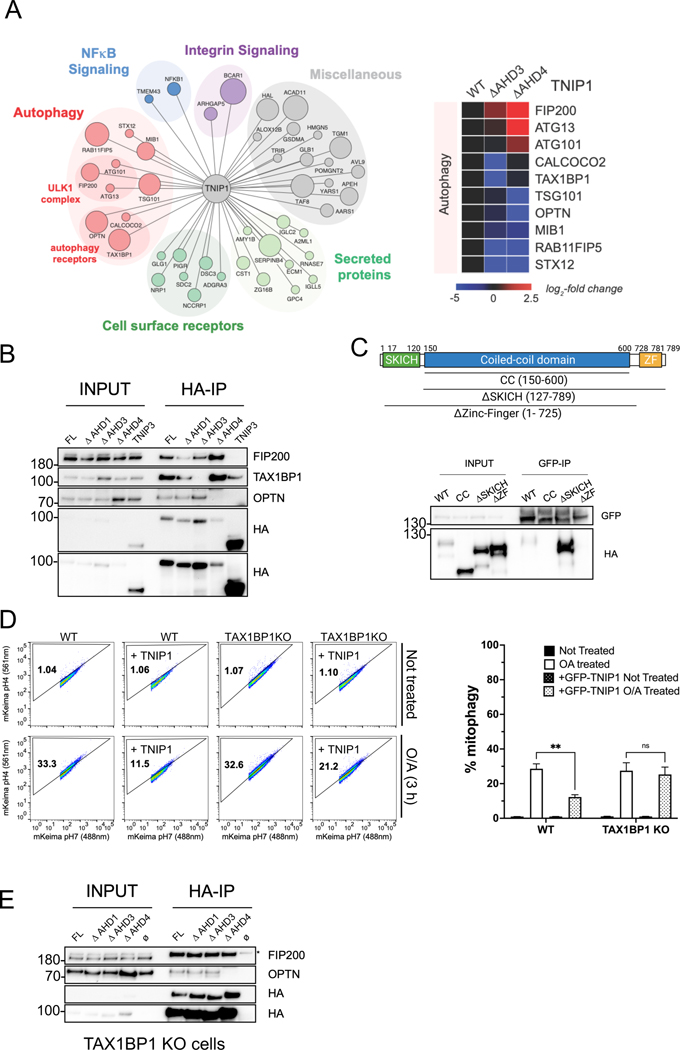
Mitophagy is a form of selective autophagy that disposes of superfluous and potentially damage-inducing organelles in a tightly controlled manner. While the machinery involved in mitophagy induction is well known, the regulation of the components is less clear. Here, we demonstrate that TNIP1 knockout in HeLa cells accelerates mitophagy rates and that ectopic TNIP1 negatively regulates the rate of mitophagy. These functions of TNIP1 depend on an evolutionarily conserved LIR motif as well as an AHD3 domain, which are required for binding to the LC3/GABARAP family of proteins and the autophagy receptor TAX1BP1, respectively. We further show that phosphorylation appears to regulate its association with the ULK1 complex member FIP200, allowing TNIP1 to compete with autophagy receptors, which provides a molecular rationale for its inhibitory function during mitophagy. Taken together, our findings describe TNIP1 as a negative regulator of mitophagy that acts at the early steps of autophagosome biogenesis.
Keywords: FIP200; FIR; LIR; Selective autophagy; TAX1BP1; TBK1; TNIP1; mitophagy; mitophagy regulation.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no financial conflicts and assure that this manuscript is original and has not been published nor is currently under consideration for publication elsewhere.
Figures
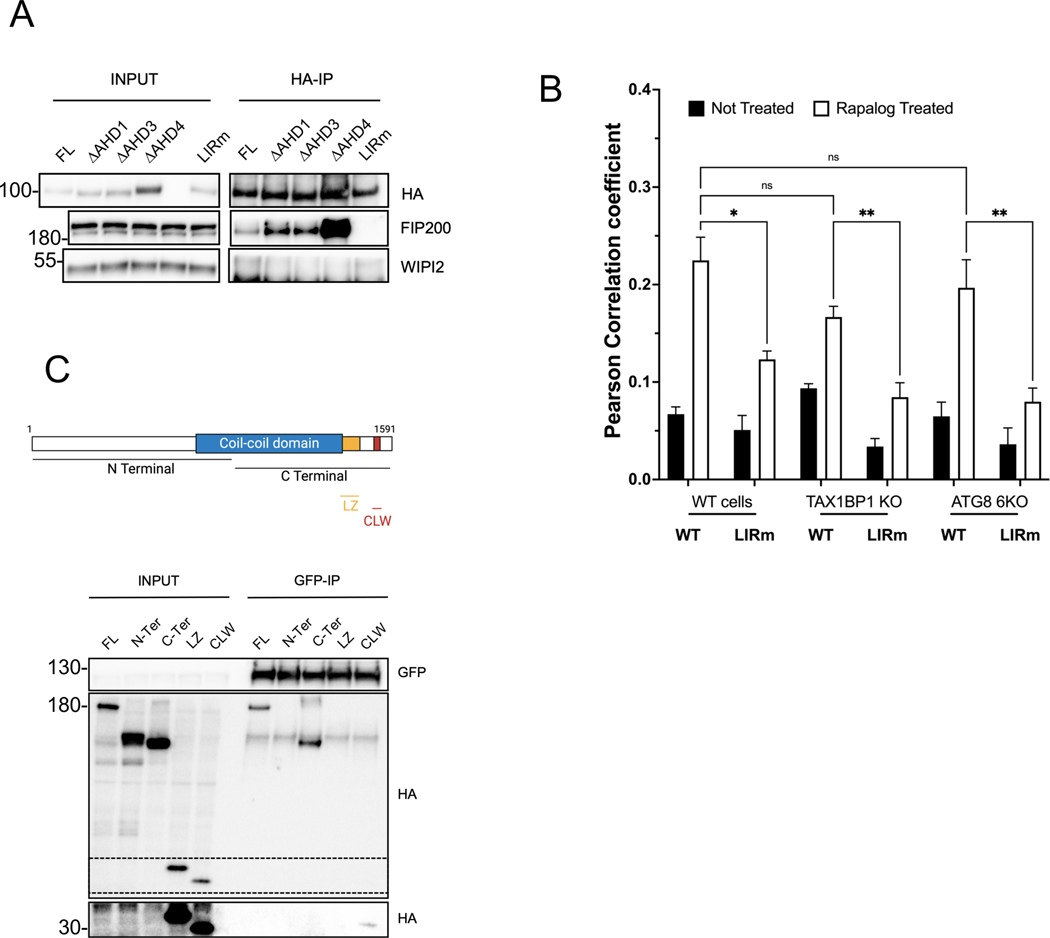
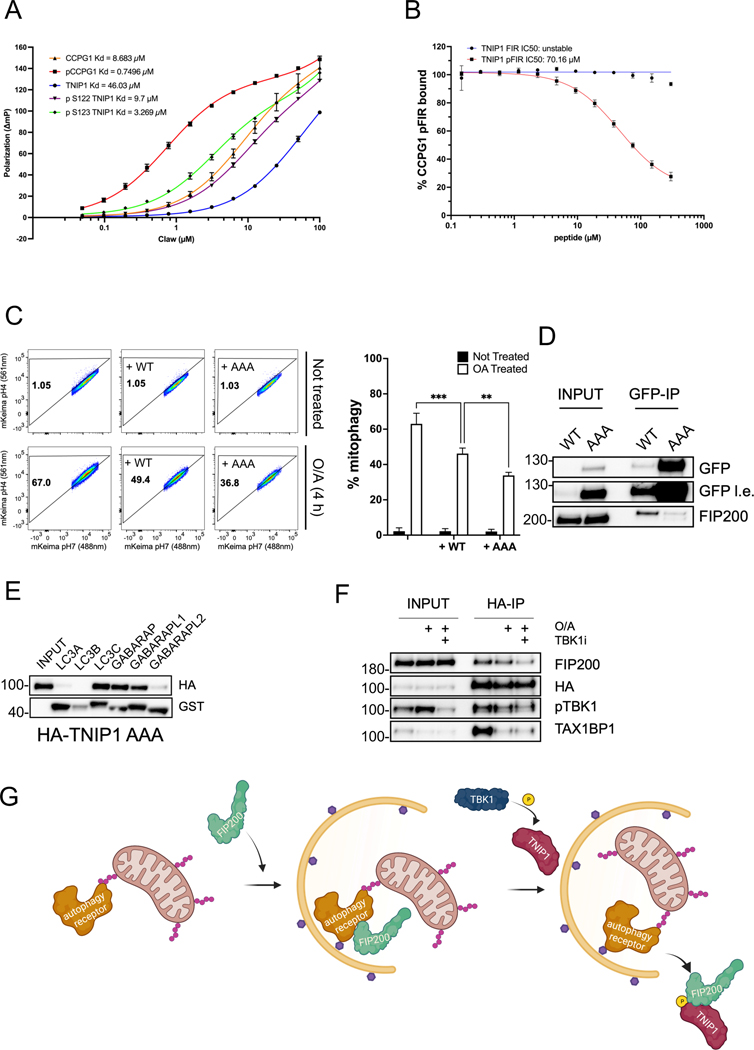
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
