

An Oxalate Transporter Gene, AtOT, Enhances Aluminum Tolerance in Arabidopsis thaliana by Regulating Oxalate Efflux
- PMID: 36901947
- PMCID: PMC10003554
- DOI: 10.3390/ijms24054516
An Oxalate Transporter Gene, AtOT, Enhances Aluminum Tolerance in Arabidopsis thaliana by Regulating Oxalate Efflux
Abstract
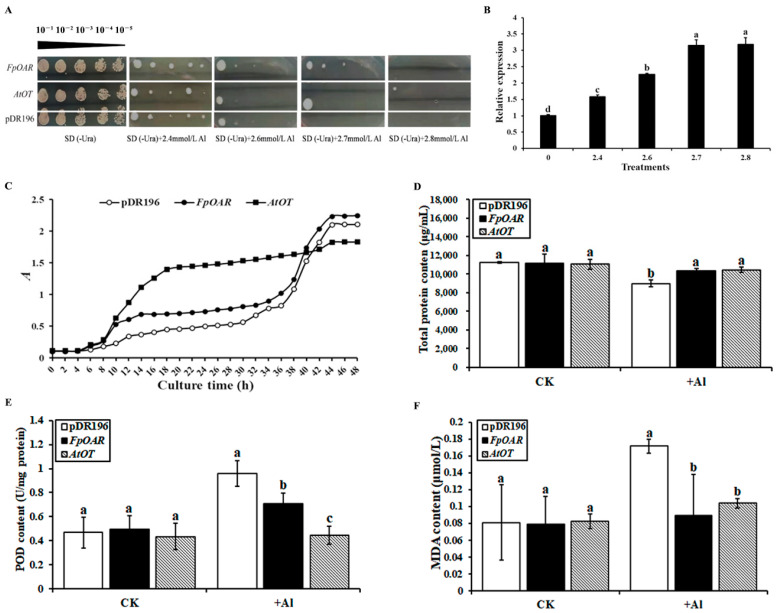
Secretion and efflux of oxalic acid from roots is an important aluminum detoxification mechanism for various plants; however, how this process is completed remains unclear. In this study, the candidate oxalate transporter gene AtOT, encoding 287 amino acids, was cloned and identified from Arabidopsis thaliana. AtOT was upregulated in response to aluminum stress at the transcriptional level, which was closely related to aluminum treatment concentration and time. The root growth of Arabidopsis was inhibited after knocking out AtOT, and this effect was amplified by aluminum stress. Yeast cells expressing AtOT enhanced oxalic acid resistance and aluminum tolerance, which was closely correlated with the secretion of oxalic acid by membrane vesicle transport. Collectively, these results underline an external exclusion mechanism of oxalate involving AtOT to enhance oxalic acid resistance and aluminum tolerance.
Keywords: Arabidopsis thaliana; AtOT; aluminum toxicity; functional characterization; gene expression; oxalate transporter.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- Silambarasan S., Logeswari P., Valentine A., Cornejo P. Role of Curtobacterium herbarum strain CAH5 on aluminum bioaccumulation and enhancement of Lactuca sativa growth under aluminum and drought stresses. Ecotoxicol. Environ. Saf. 2019;183:109573.1–109573.10. doi: 10.1016/j.ecoenv.2019.109573. - DOI - PubMed
-
- Feng Y.M., Yu M., Wang C.Q., Liu J.Y. Aluminum toxicity induced cell responses in higher plants. J. Huazhong Agric. Univ. 2005;24:320–324.
-
- Zhang H., Zhang G.L., Zhao Y.G., Zhao W.J., Qi Z.P. Chemical degradation of a Ferralsol (Oxisol) under intensive rubber (Hevea Brasiliensis) farming in tropical China. Soil Tillage Res. 2007;93:109–116. doi: 10.1016/j.still.2006.03.013. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
