

Molecular mechanism of biased signaling at the kappa opioid receptor
- PMID: 36906681
- PMCID: PMC10008561
- DOI: 10.1038/s41467-023-37041-7
Molecular mechanism of biased signaling at the kappa opioid receptor
Abstract
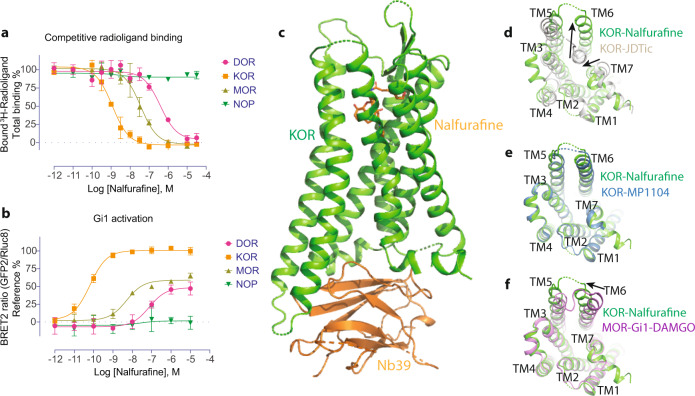
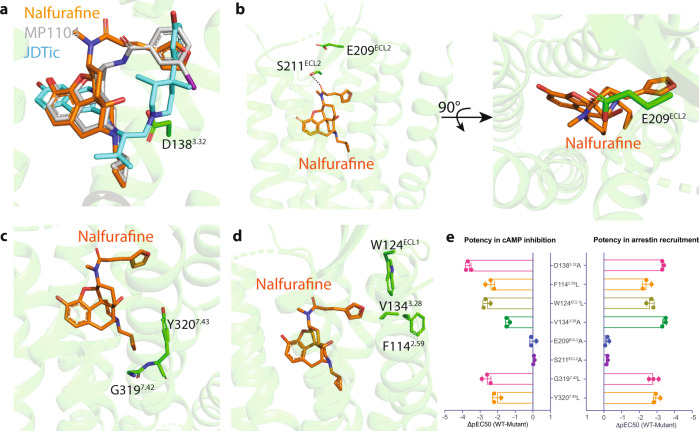
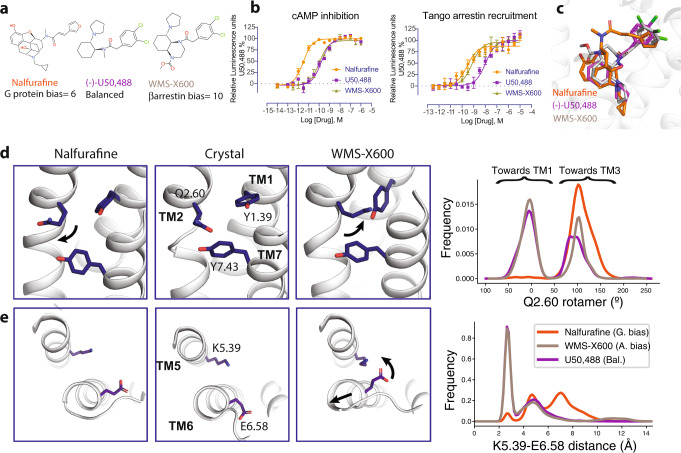
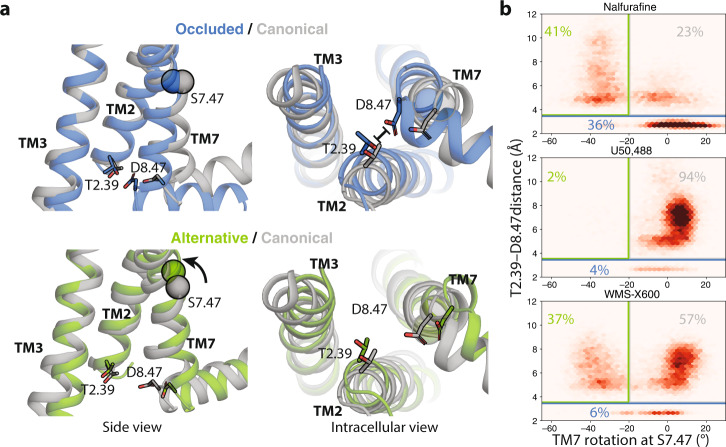
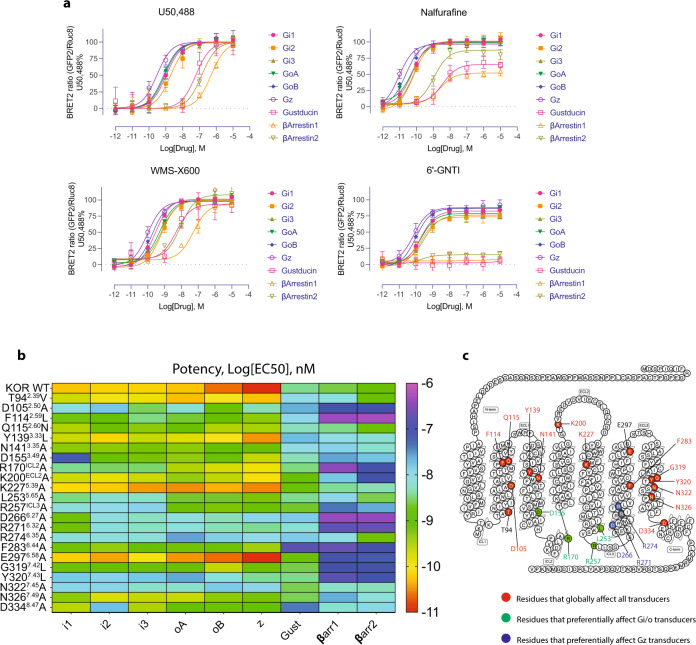
The κ-opioid receptor (KOR) has emerged as an attractive drug target for pain management without addiction, and biased signaling through particular pathways of KOR may be key to maintaining this benefit while minimizing side-effect liabilities. As for most G protein-coupled receptors (GPCRs), however, the molecular mechanisms of ligand-specific signaling at KOR have remained unclear. To better understand the molecular determinants of KOR signaling bias, we apply structure determination, atomic-level molecular dynamics (MD) simulations, and functional assays. We determine a crystal structure of KOR bound to the G protein-biased agonist nalfurafine, the first approved KOR-targeting drug. We also identify an arrestin-biased KOR agonist, WMS-X600. Using MD simulations of KOR bound to nalfurafine, WMS-X600, and a balanced agonist U50,488, we identify three active-state receptor conformations, including one that appears to favor arrestin signaling over G protein signaling and another that appears to favor G protein signaling over arrestin signaling. These results, combined with mutagenesis validation, provide a molecular explanation of how agonists achieve biased signaling at KOR.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
