

This is a preprint.
A scalable, GMP-compatible, autologous organotypic cell therapy for Dystrophic Epidermolysis Bullosa
- PMID: 36909618
- PMCID: PMC10002612
- DOI: 10.1101/2023.02.28.529447
A scalable, GMP-compatible, autologous organotypic cell therapy for Dystrophic Epidermolysis Bullosa
Update in
-
A scalable and cGMP-compatible autologous organotypic cell therapy for Dystrophic Epidermolysis Bullosa.Nat Commun. 2024 Jul 11;15(1):5834. doi: 10.1038/s41467-024-49400-z. Nat Commun. 2024. PMID: 38992003 Free PMC article.
Abstract
Background: Gene editing in induced pluripotent stem (iPS) cells has been hailed to enable new cell therapies for various monogenetic diseases including dystrophic epidermolysis bullosa (DEB). However, manufacturing, efficacy and safety roadblocks have limited the development of genetically corrected, autologous iPS cell-based therapies.
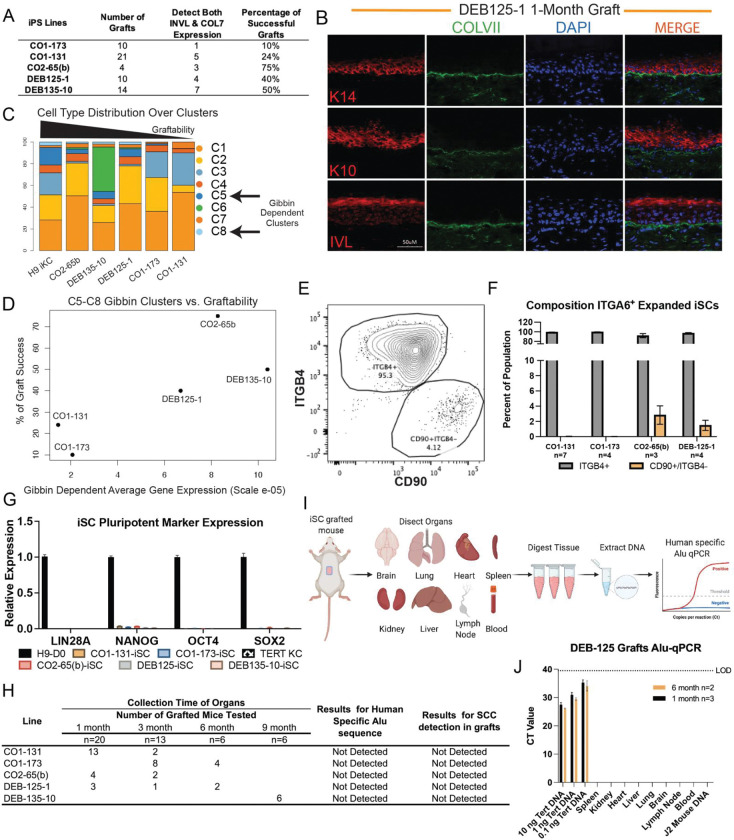
Methods: We developed Dystrophic Epidermolysis Bullosa Cell Therapy (DEBCT), a new generation GMP-compatible (cGMP), reproducible, and scalable platform to produce autologous clinical-grade iPS cell-derived organotypic induced skin composite (iSC) grafts to treat incurable wounds of patients lacking type VII collagen (C7). DEBCT uses a combined high-efficiency reprogramming and CRISPR-based genetic correction single step to generate genome scar-free, COL7A1 corrected clonal iPS cells from primary patient fibroblasts. Validated iPS cells are converted into epidermal, dermal and melanocyte progenitors with a novel 2D organoid differentiation protocol, followed by CD49f enrichment and expansion to minimize maturation heterogeneity. iSC product characterization by single cell transcriptomics was followed by mouse xenografting for disease correcting activity at 1 month and toxicology analysis at 1-6 months. Culture-acquired mutations, potential CRISPR-off targets, and cancer-driver variants were evaluated by targeted and whole genome sequencing.
Findings: iPS cell-derived iSC grafts were reproducibly generated from four recessive DEB patients with different pathogenic mutations. Organotypic iSC grafts onto immune-compromised mice developed into stable stratified skin with functional C7 restoration. Single cell transcriptomic characterization of iSCs revealed prominent holoclone stem cell signatures in keratinocytes and the recently described Gibbin-dependent signature in dermal fibroblasts. The latter correlated with enhanced graftability. Multiple orthogonal sequencing and subsequent computational approaches identified random and non-oncogenic mutations introduced by the manufacturing process. Toxicology revealed no detectable tumors after 3-6 months in DEBCT-treated mice.
Interpretation: DEBCT successfully overcomes previous roadblocks and represents a robust, scalable, and safe cGMP manufacturing platform for production of a CRISPR-corrected autologous organotypic skin graft to heal DEB patient wounds.
Keywords: CRISPR-CAS9; Epidermolysis Bullosa; Induced Pluripotent Stem Cells (iPS cells); dermal fibroblasts; epithelium; gene editing; good manufacturing practices compatibility (c-GMP); keratinocyte; skin; tissue engineering; wound healing.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no competing or conflicting interests.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous