

Antisense oligonucleotide therapy corrects splicing in the common Stargardt disease type 1-causing variant ABCA4 c.5461-10T>C
- PMID: 36910710
- PMCID: PMC9999166
- DOI: 10.1016/j.omtn.2023.02.020
Antisense oligonucleotide therapy corrects splicing in the common Stargardt disease type 1-causing variant ABCA4 c.5461-10T>C
Abstract
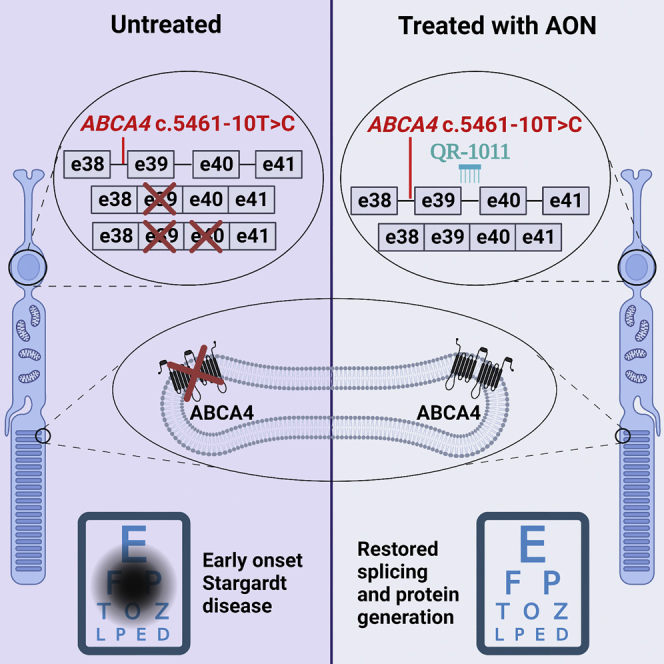
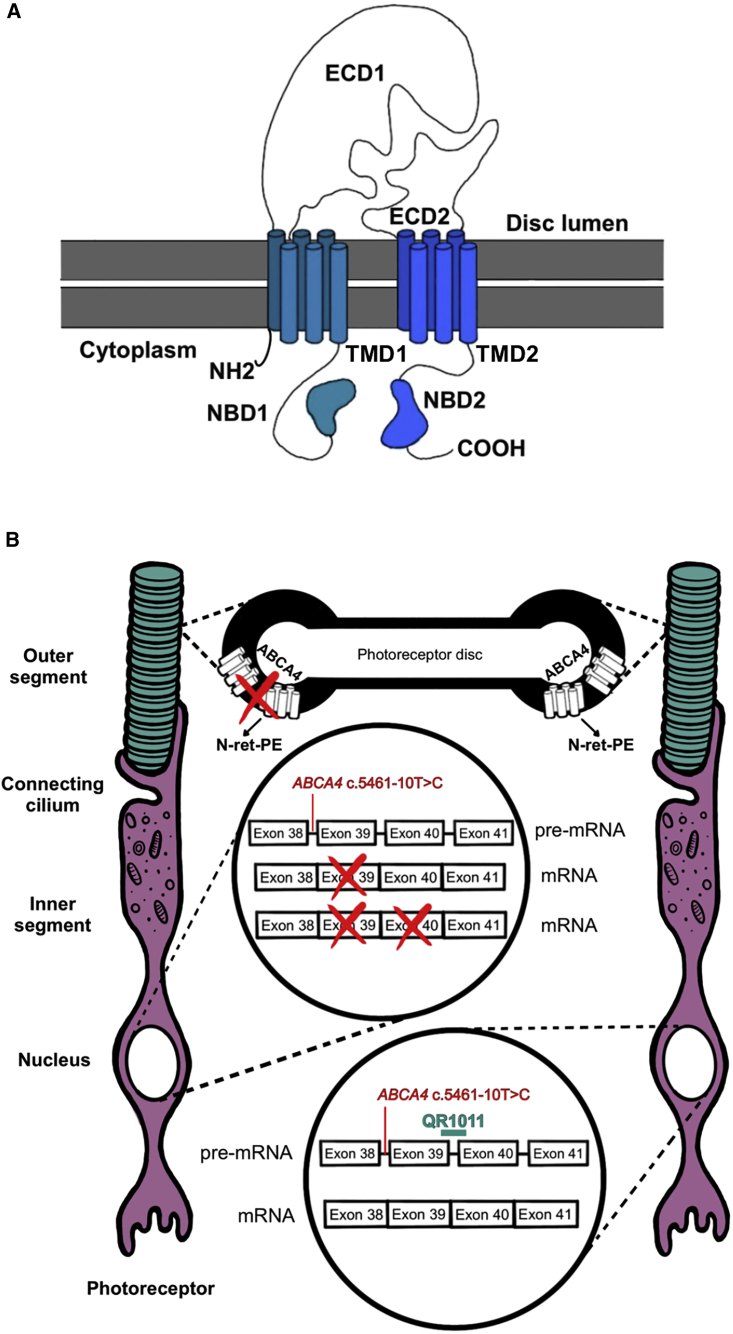
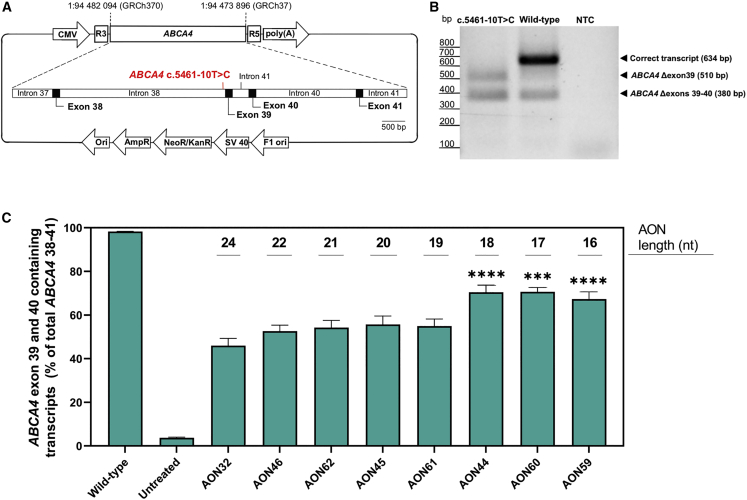
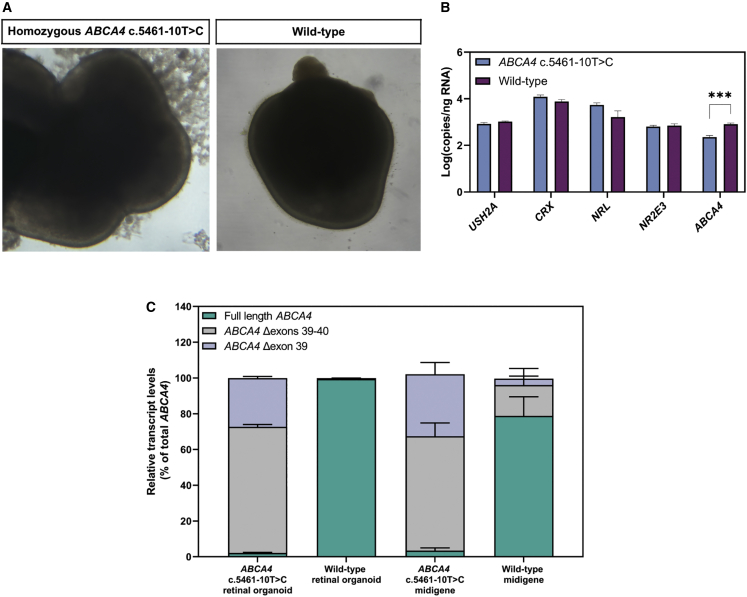
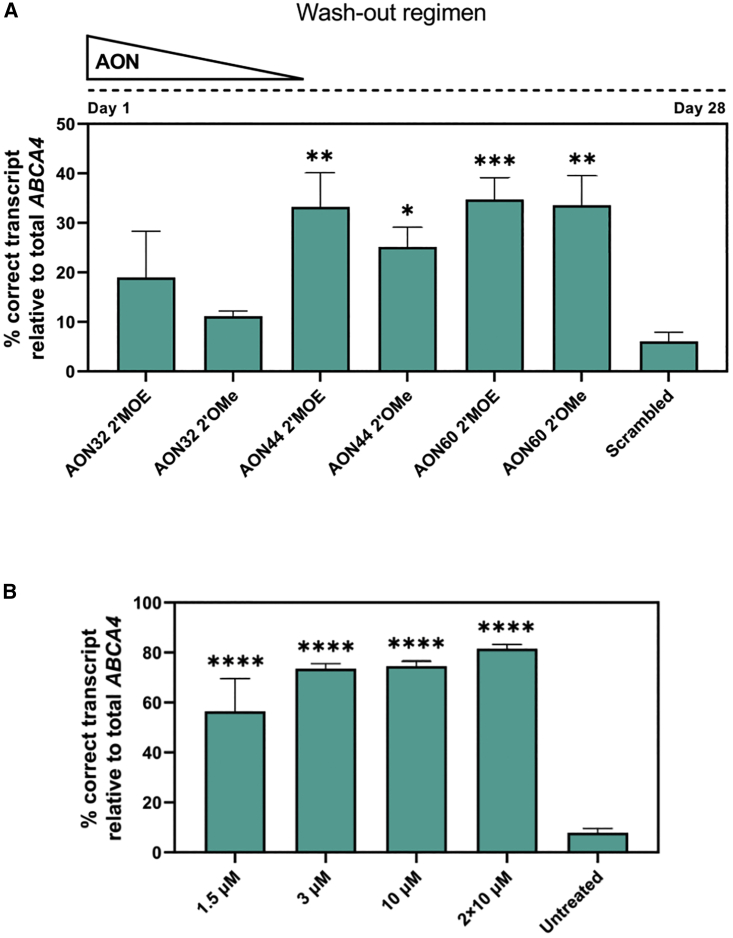
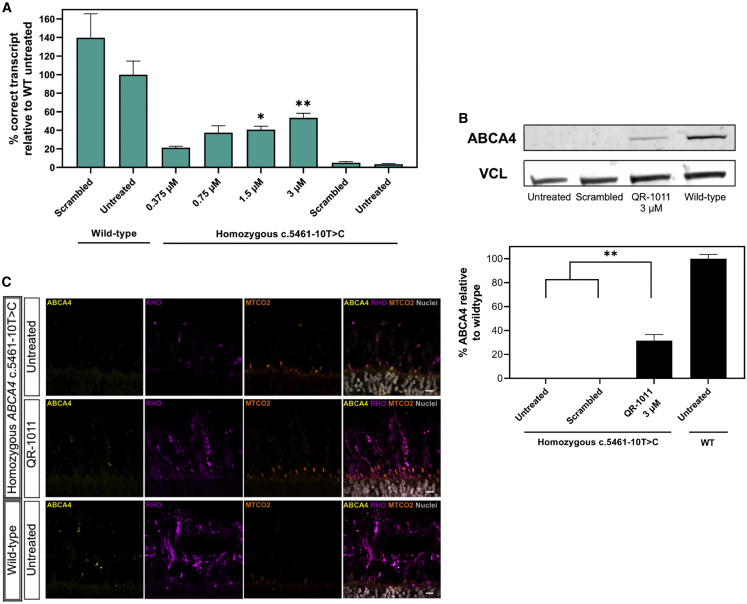
Stargardt disease type 1 (STGD1) is the most common hereditary form of maculopathy and remains untreatable. STGD1 is caused by biallelic variants in the ABCA4 gene, which encodes the ATP-binding cassette (type 4) protein (ABCA4) that clears toxic byproducts of the visual cycle. The c.5461-10T>C p.[Thr1821Aspfs∗6,Thr1821Valfs∗13] variant is the most common severe disease-associated variant, and leads to exon skipping and out-of-frame ABCA4 transcripts that prevent translation of functional ABCA4 protein. Homozygous individuals typically display early onset STGD1 and are legally blind by early adulthood. Here, we applied antisense oligonucleotides (AONs) to promote exon inclusion and restore wild-type RNA splicing of ABCA4 c.5461-10T>C. The effect of AONs was first investigated in vitro using an ABCA4 midigene model. Subsequently, the best performing AONs were administered to homozygous c.5461-10T>C 3D human retinal organoids. Isoform-specific digital polymerase chain reaction revealed a significant increase in correctly spliced transcripts after treatment with the lead AON, QR-1011, up to 53% correct transcripts at a 3 μM dose. Furthermore, western blot and immunohistochemistry analyses identified restoration of ABCA4 protein after treatment. Collectively, we identified QR-1011 as a potent splice-correcting AON and a possible therapeutic intervention for patients harboring the severe ABCA4 c.5461-10T>C variant.
Keywords: ABCA4; MT: Oligonucleotides: Therapies and Applications; RNA therapy; Stargardt disease; antisense oligonucleotides; retinal organoids; splicing correction.
© 2023 The Authors.
Conflict of interest statement
M.K., P.d.B., K.D., T.H., W.B., G.P., and J.S. were employed by ProQR Therapeutics during this project. K.D. is inventor on the international patent application (WO2018189376) that has been filed by ProQR Therapeutics describing methods and means for AON therapy for Stargardt disease type 1. The rest of the co-authors declare that they have no competing interests.
Figures
References
-
- Allikmets R., Singh N., Sun H., Shroyer N.F., Hutchinson A., Chidambaram A., Gerrard B., Baird L., Stauffer D., Peiffer A., et al. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat. Nat. Genet. 1997;15:236–246. doi: 10.1038/ng0397-236. - DOI - PubMed
-
- Cremers F.P., van de Pol D.J., van Driel M., den Hollander A.I., van Haren F.J., Knoers N.V., Tijmes N., Bergen A.A., Rohrschneider K., Blankenagel A., et al. Autosomal recessive retinitis pigmentosa and cone-rod dystrophy caused by splice site mutations in the Stargardt's disease gene ABCR. Hum. Mol. Genet. 1998;7:355–362. doi: 10.1093/hmg/7.3.355. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
