

The Effect of Developmental Pleiotropy on the Evolution of Insect Immune Genes
- PMID: 36911982
- PMCID: PMC10063218
- DOI: 10.1093/gbe/evad044
The Effect of Developmental Pleiotropy on the Evolution of Insect Immune Genes
Abstract
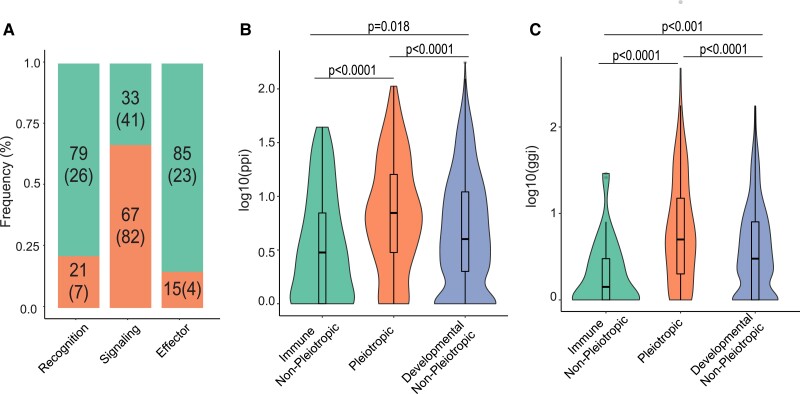
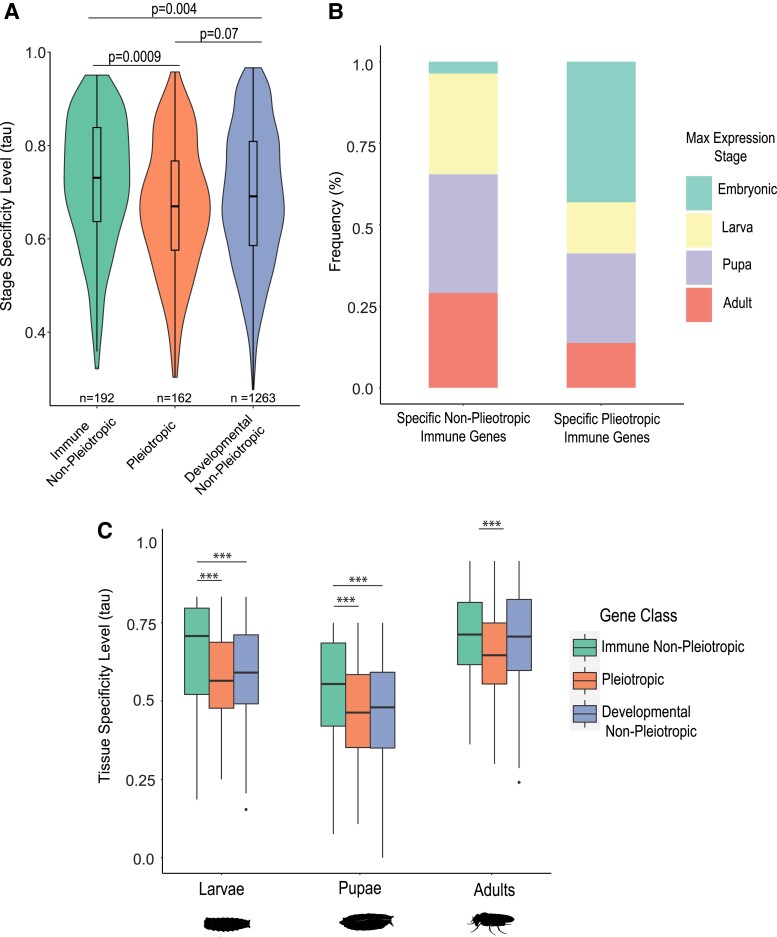
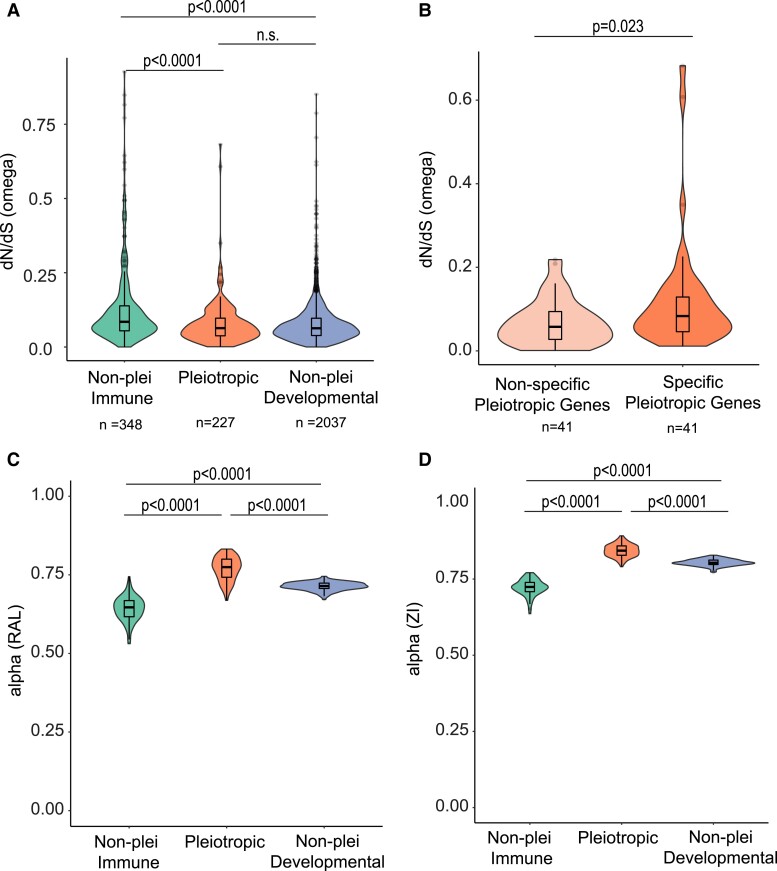
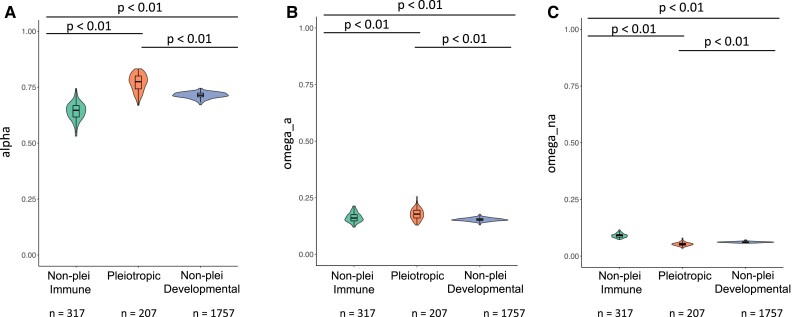
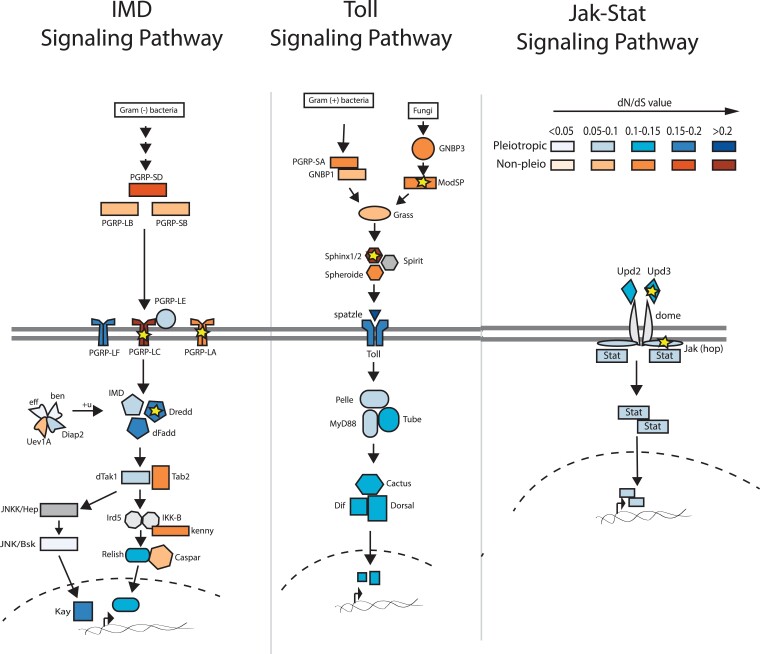
The pressure to survive ever-changing pathogen exposure explains the frequent observation that immune genes are among the fastest evolving in the genomes of many taxa, but an intriguing proportion of immune genes also appear to be under purifying selection. Though variance in evolutionary signatures of immune genes is often attributed to differences in gene-specific interactions with microbes, this explanation neglects the possibility that immune genes participate in other biological processes that could pleiotropically constrain adaptive selection. In this study, we analyzed available transcriptomic and genomic data from Drosophila melanogaster and related species to test the hypothesis that there is substantial pleiotropic overlap in the developmental and immunological functions of genes involved in immune signaling and that pleiotropy would be associated with stronger signatures of evolutionary constraint. Our results suggest that pleiotropic immune genes do evolve more slowly than those having no known developmental functions and that signatures of constraint are particularly strong for pleiotropic immune genes that are broadly expressed across life stages. These results support the general yet untested hypothesis that pleiotropy can constrain immune system evolution, raising new fundamental questions about the benefits of maintaining pleiotropy in systems that need to rapidly adapt to changing pathogen pressures.
Keywords: Toll pathway; adaptive evolution; evolutionary constraint; insect immunity; molecular evolution.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures
References
-
- Anthoney N, Foldi I, Hidalgo A. 2018. Toll and Toll-like receptor signalling in development. Development 145:dev156018. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
