

Cerebrospinal Fluid and Brain Proteoforms of the Granin Neuropeptide Family in Alzheimer's Disease
- PMID: 36912488
- PMCID: PMC10080684
- DOI: 10.1021/jasms.2c00341
Cerebrospinal Fluid and Brain Proteoforms of the Granin Neuropeptide Family in Alzheimer's Disease
Erratum in
-
Correction to "Cerebrospinal Fluid and Brain Proteoforms of the Granin Neuropeptide Family in Alzheimer's Disease".J Am Soc Mass Spectrom. 2023 Oct 4;34(10):2435-2436. doi: 10.1021/jasms.3c00265. Epub 2023 Aug 15. J Am Soc Mass Spectrom. 2023. PMID: 37581263 Free PMC article. No abstract available.
Abstract
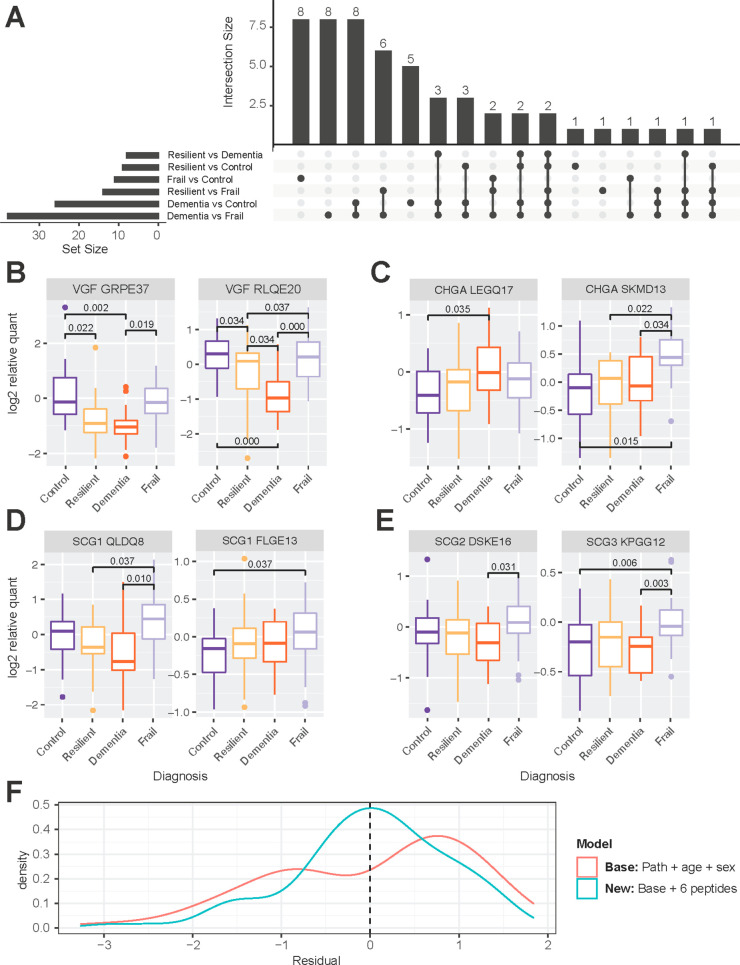
The granin neuropeptide family is composed of acidic secretory signaling molecules that act throughout the nervous system to help modulate synaptic signaling and neural activity. Granin neuropeptides have been shown to be dysregulated in different forms of dementia, including Alzheimer's disease (AD). Recent studies have suggested that the granin neuropeptides and their protease-cleaved bioactive peptides (proteoforms) may act as both powerful drivers of gene expression and as a biomarker of synaptic health in AD. The complexity of granin proteoforms in human cerebrospinal fluid (CSF) and brain tissue has not been directly addressed. We developed a reliable nontryptic mass spectrometry assay to comprehensively map and quantify endogenous neuropeptide proteoforms in the brain and CSF of individuals diagnosed with mild cognitive impairment and dementia due to AD compared to healthy controls, individuals with preserved cognition despite AD pathology ("Resilient"), and those with impaired cognition but no AD or other discernible pathology ("Frail"). We drew associations between neuropeptide proteoforms, cognitive status, and AD pathology values. Decreased levels of VGF proteoforms were observed in CSF and brain tissue from individuals with AD compared to controls, while select proteoforms from chromogranin A showed the opposite effect. To address mechanisms of neuropeptide proteoform regulation, we showed that the proteases Calpain-1 and Cathepsin S can cleave chromogranin A, secretogranin-1, and VGF into proteoforms found in both the brain and CSF. We were unable to demonstrate differences in protease abundance in protein extracts from matched brains, suggesting that regulation may occur at the level of transcription.
Conflict of interest statement
The authors declare the following competing financial interest(s): S. Arnold has received honoraria and/or travel expenses for lectures from Abbvie, Eisai, and Biogen and has served on scientifc advisory boards of Corte, has received consulting fees from Athira, Cassava, Cognito Therapeutics, EIP Pharma and Orthogonal Neuroscience, and has received research grant support from NIH, Alzheimers Association, Alzheimers Drug Discovery Foundation, Abbvie, Amylyx, EIP Pharma, Merck, Janssen/Johnson & Johnson, Novartis, and vTv.
Figures
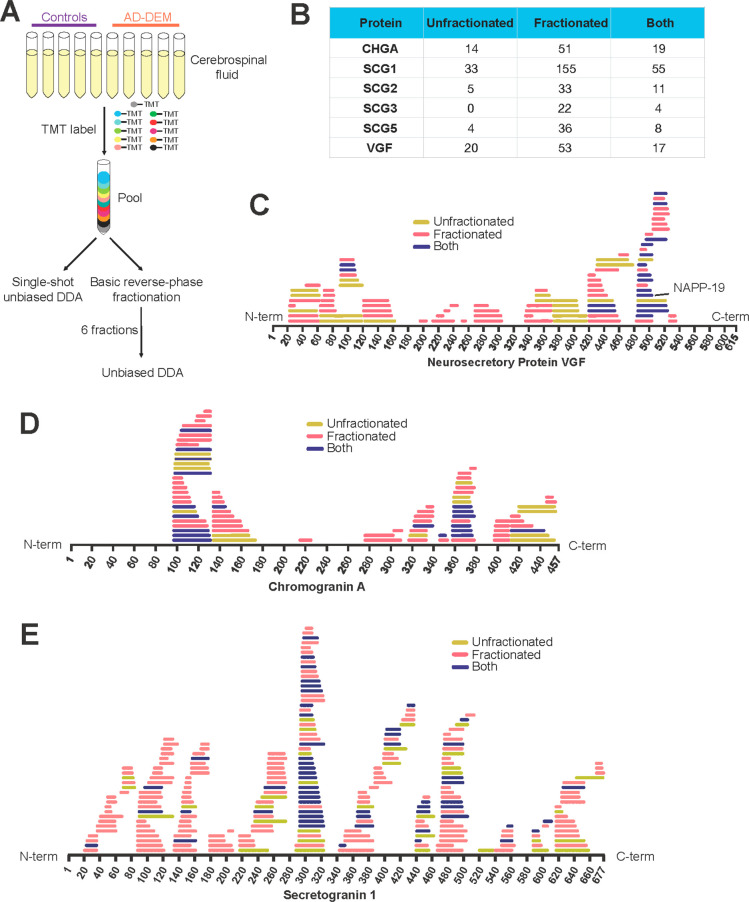
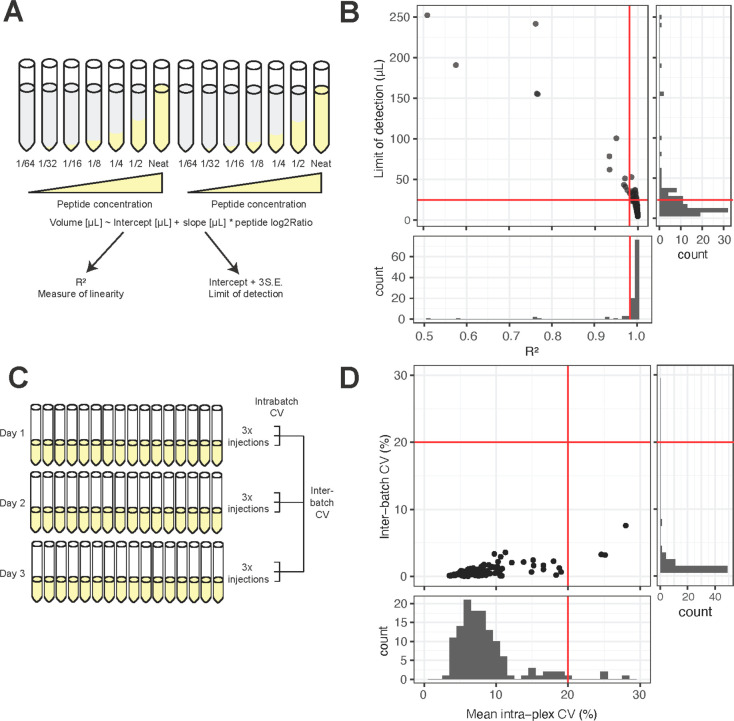
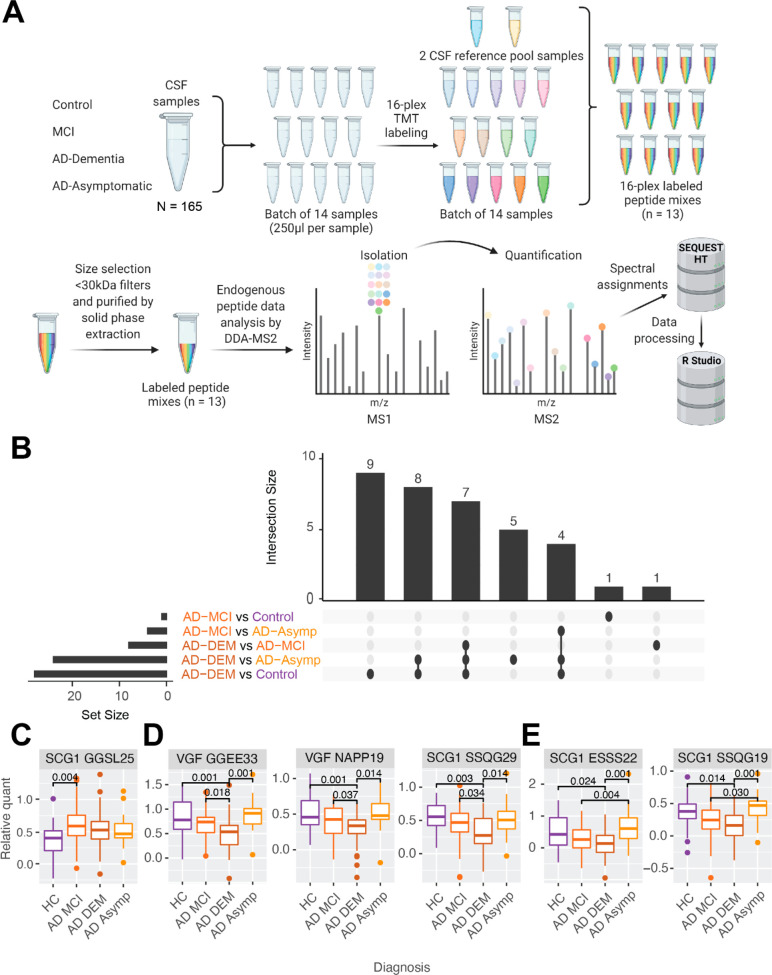
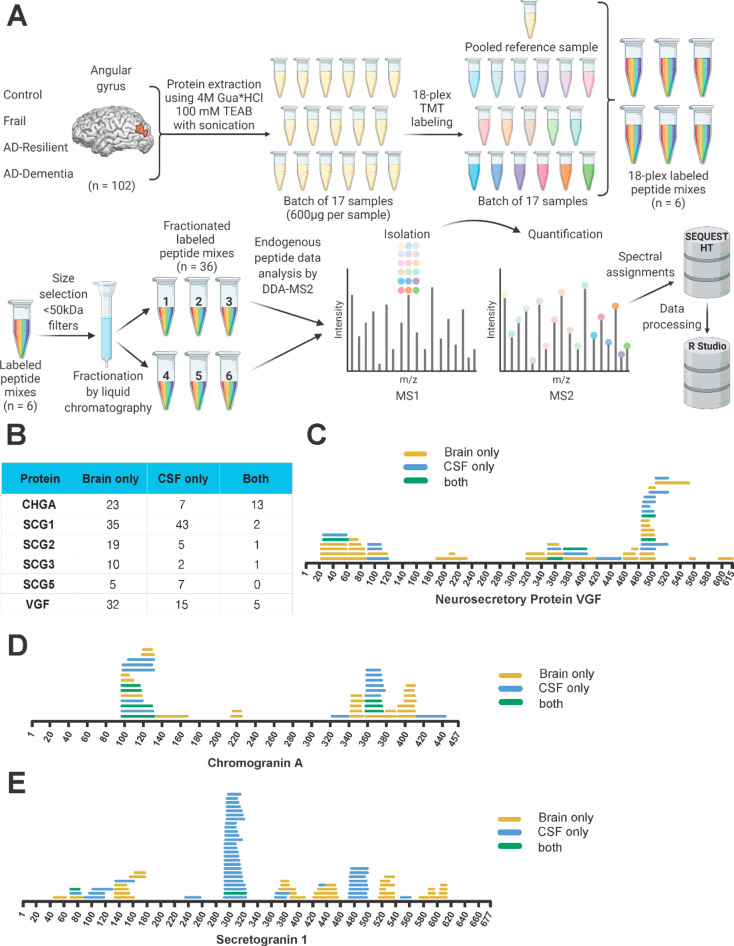



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
