

Meisosomes, folded membrane microdomains between the apical extracellular matrix and epidermis
- PMID: 36913486
- PMCID: PMC10010689
- DOI: 10.7554/eLife.75906
Meisosomes, folded membrane microdomains between the apical extracellular matrix and epidermis
Abstract
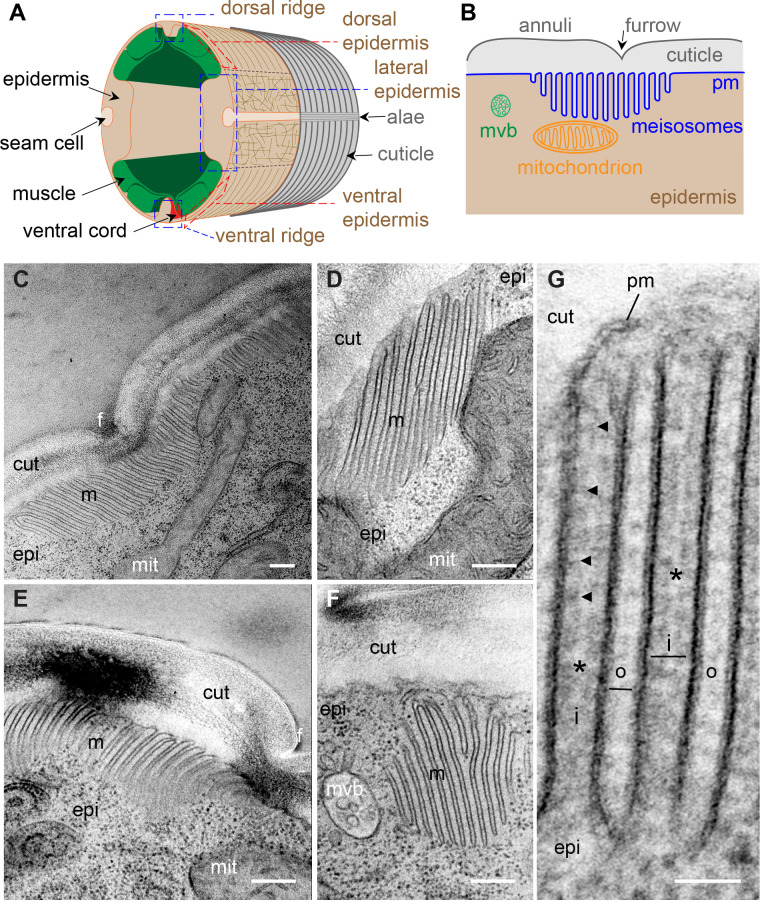
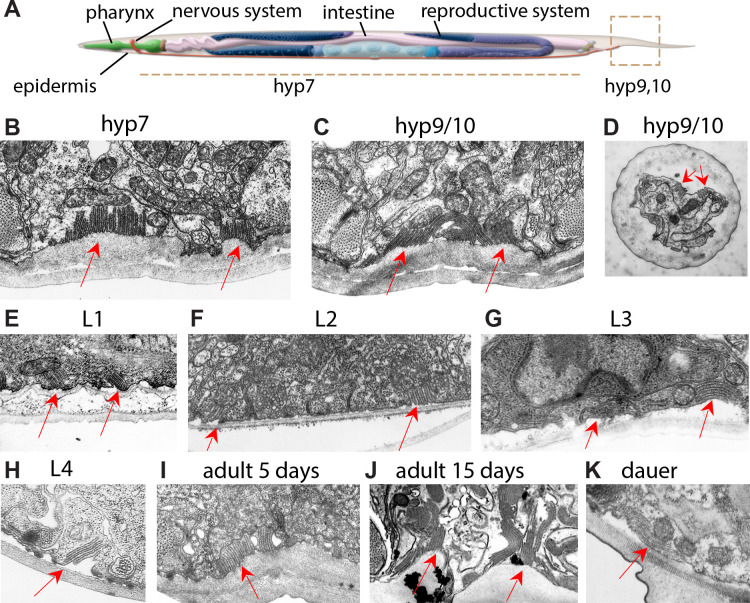
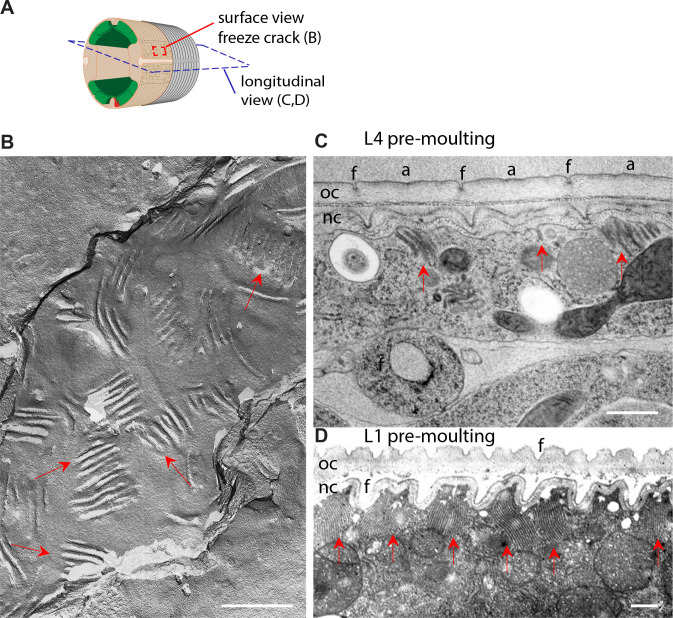
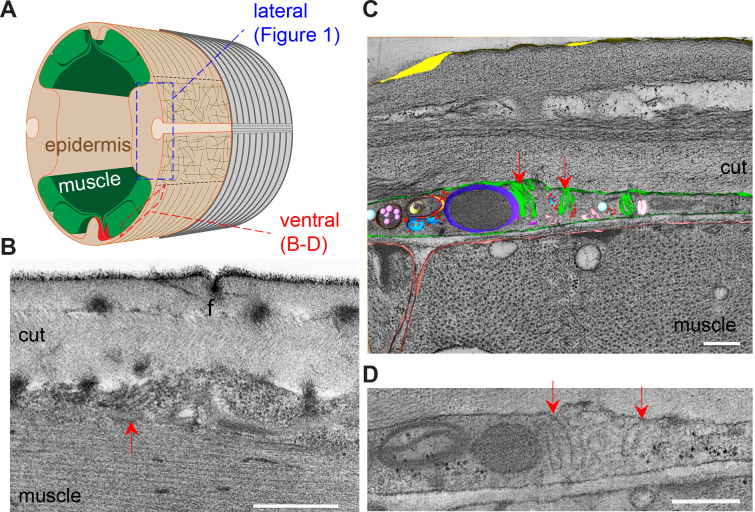
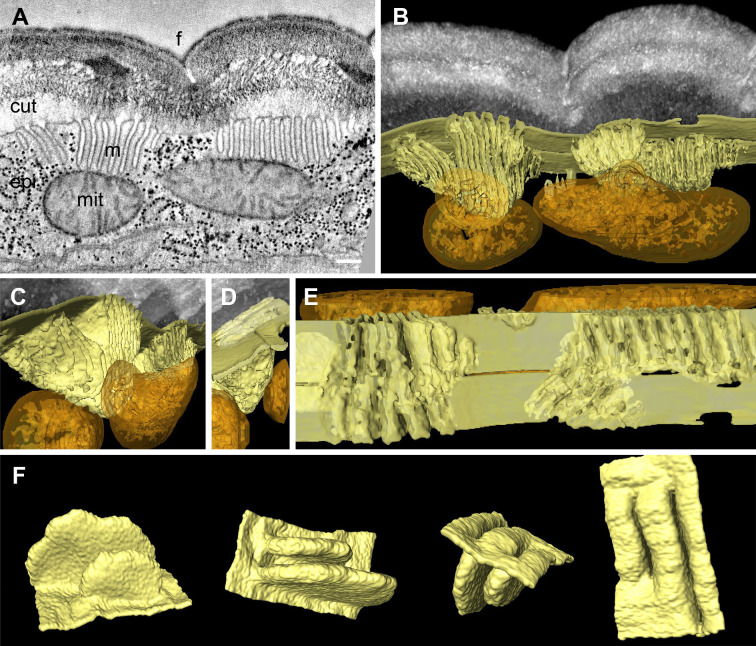
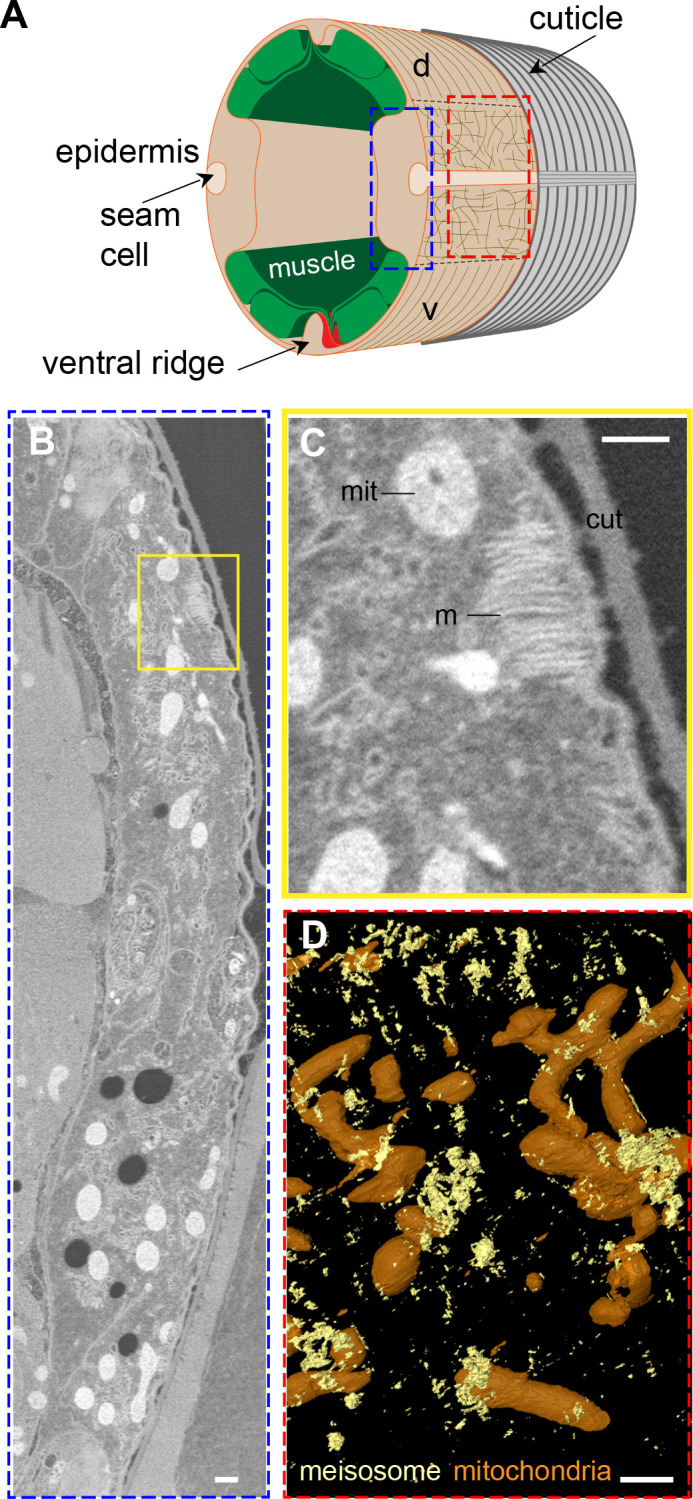
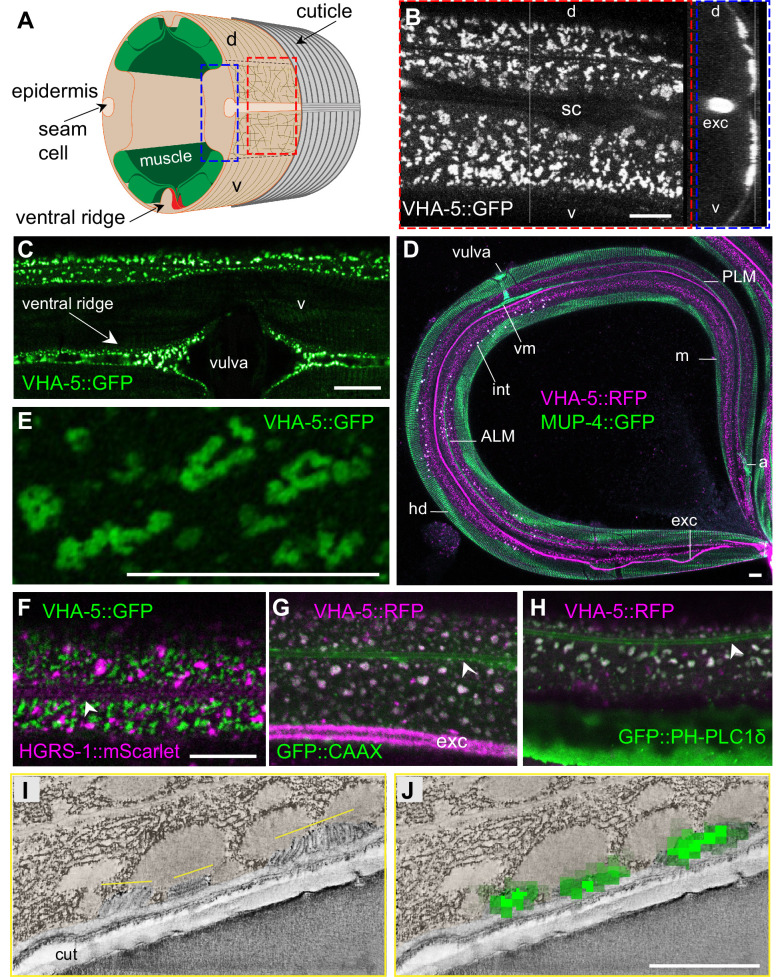
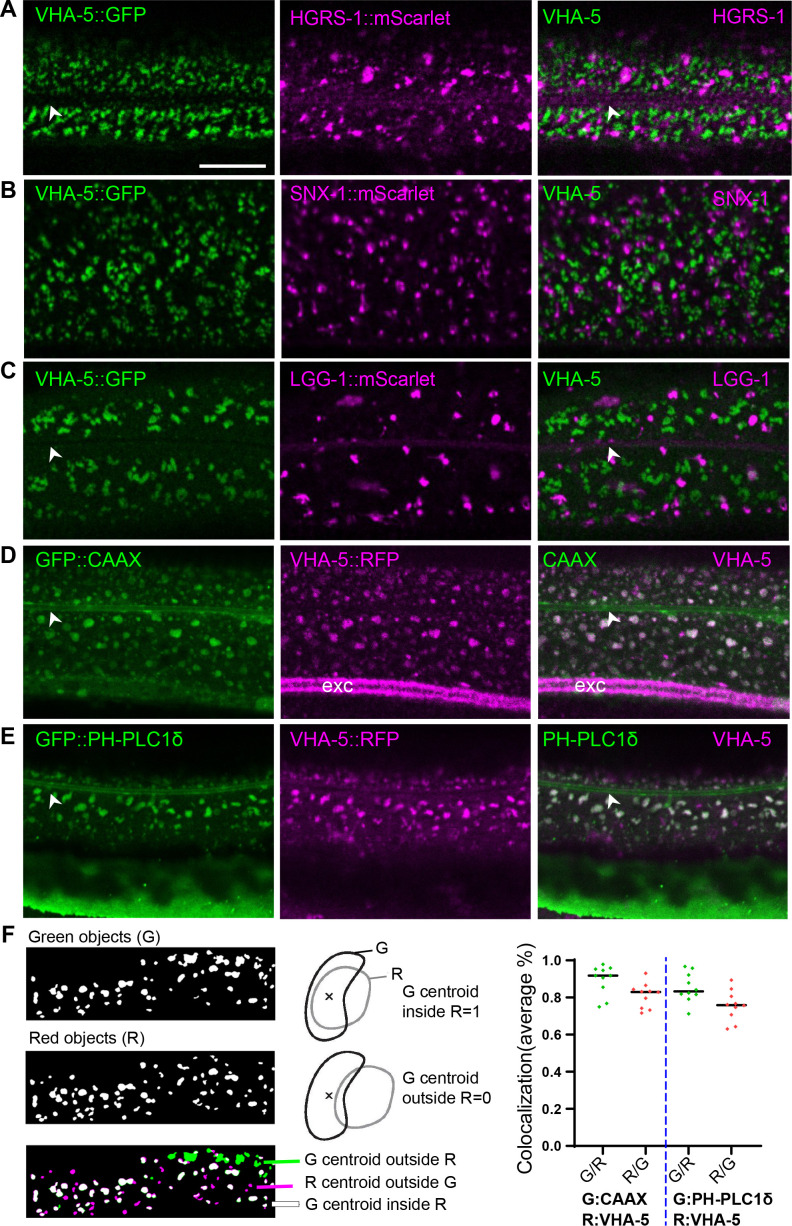
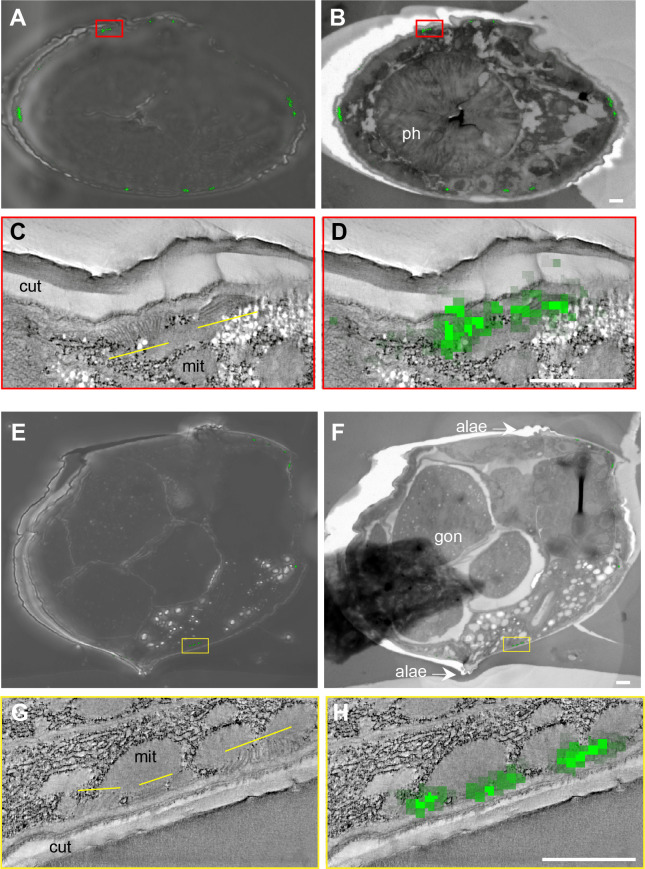
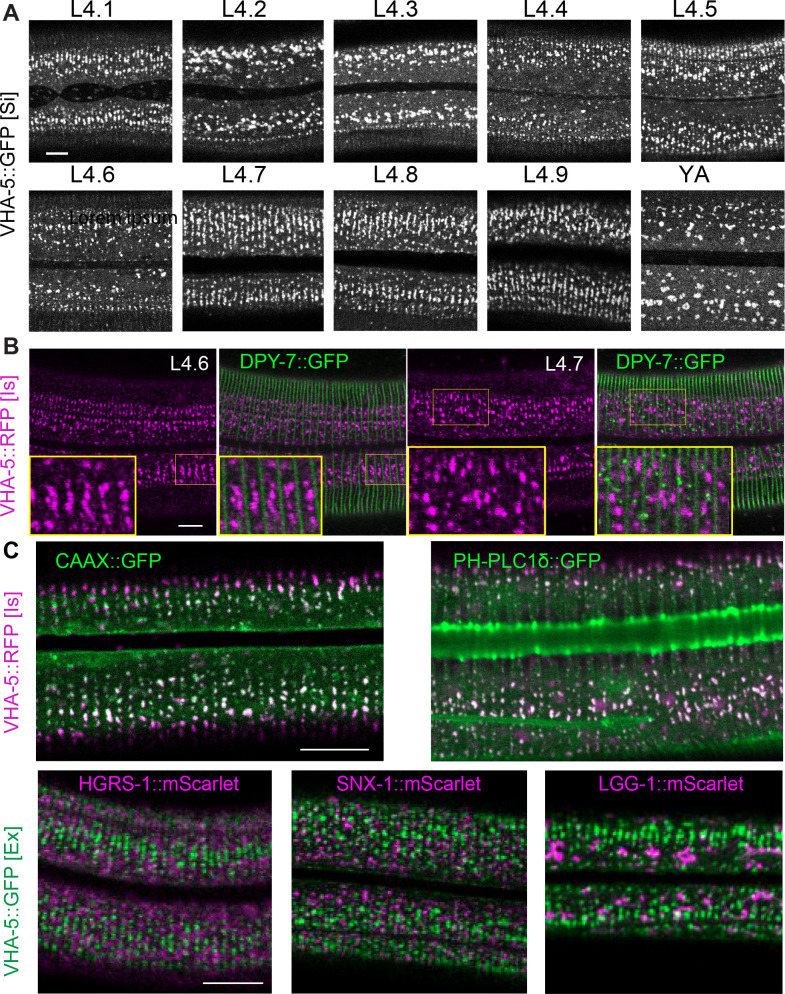
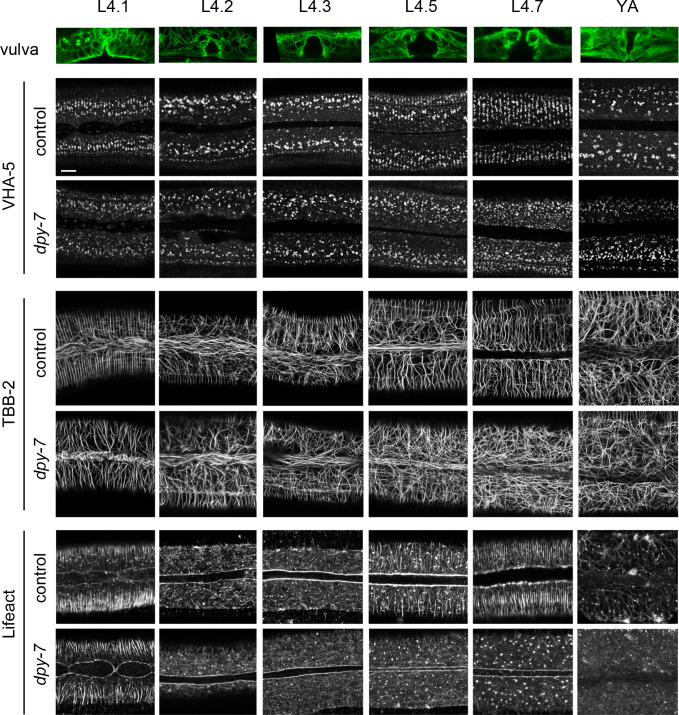
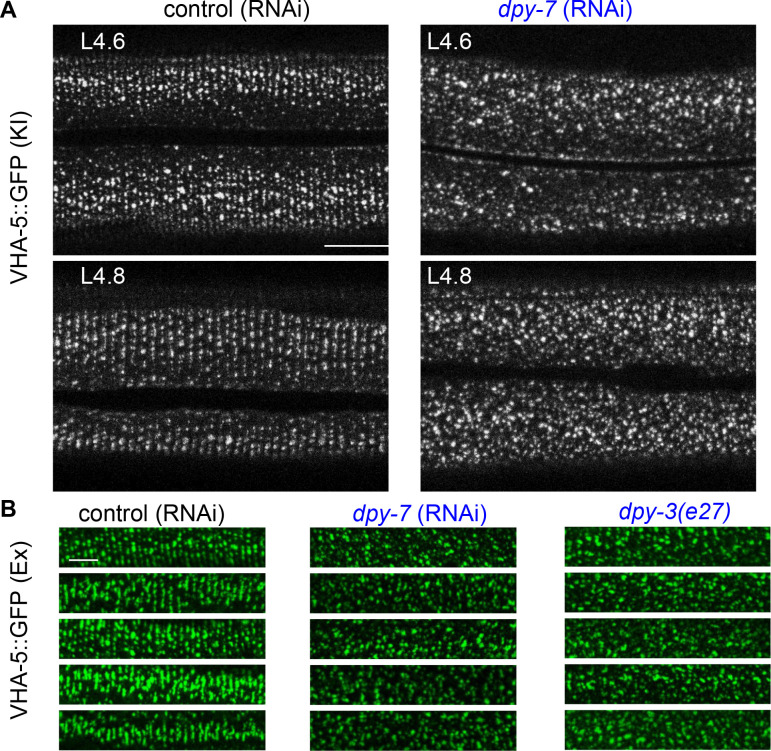
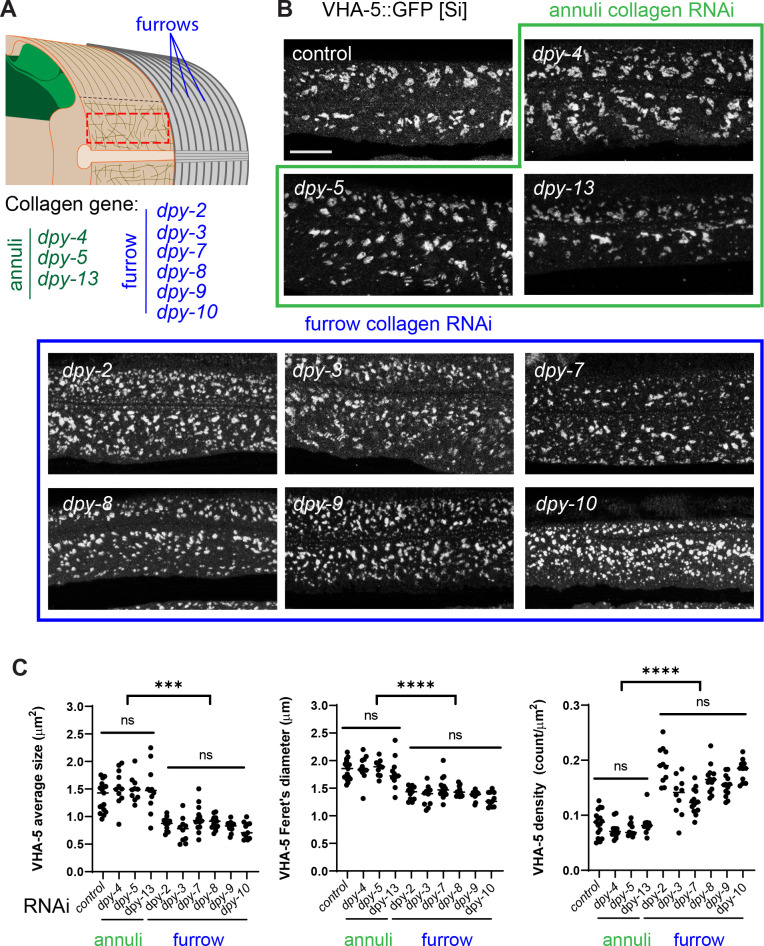
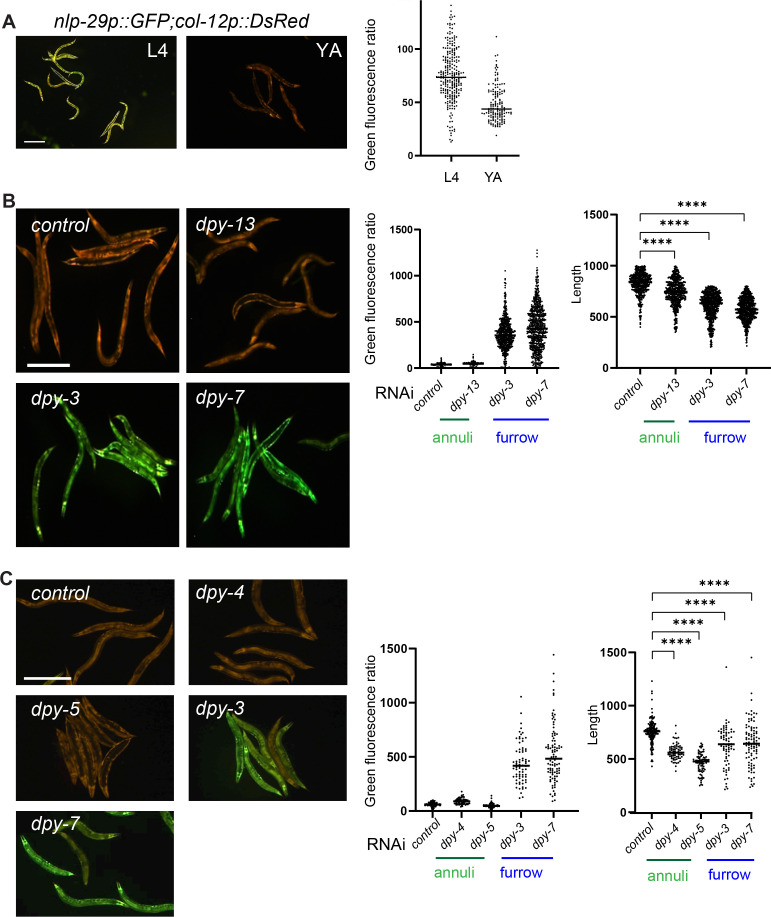
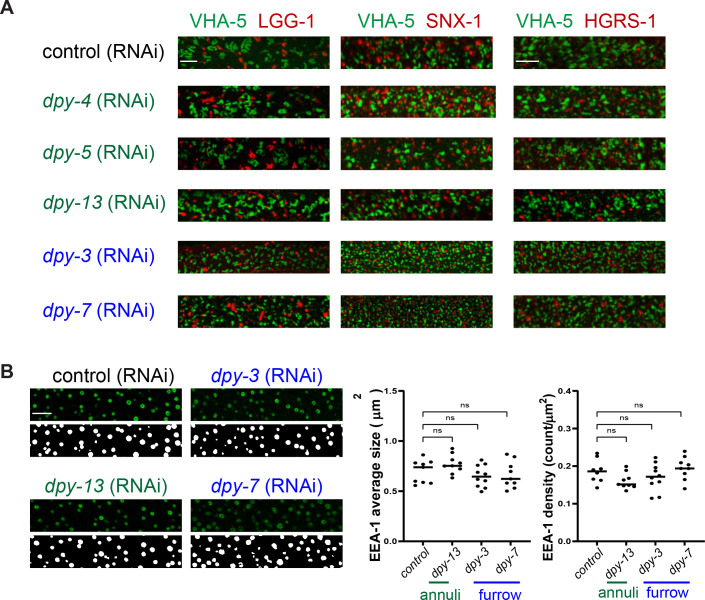
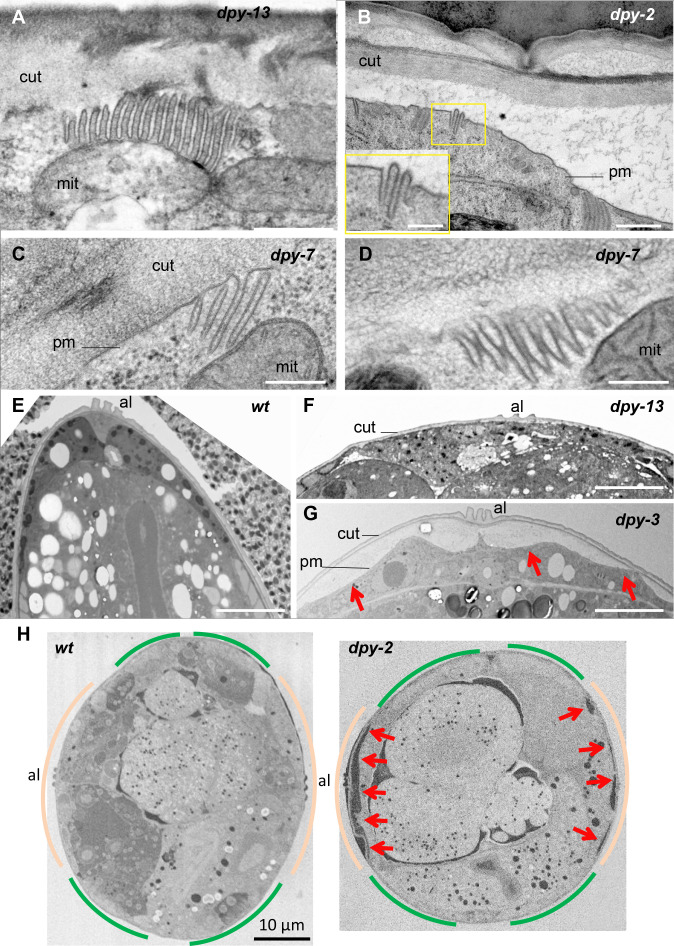
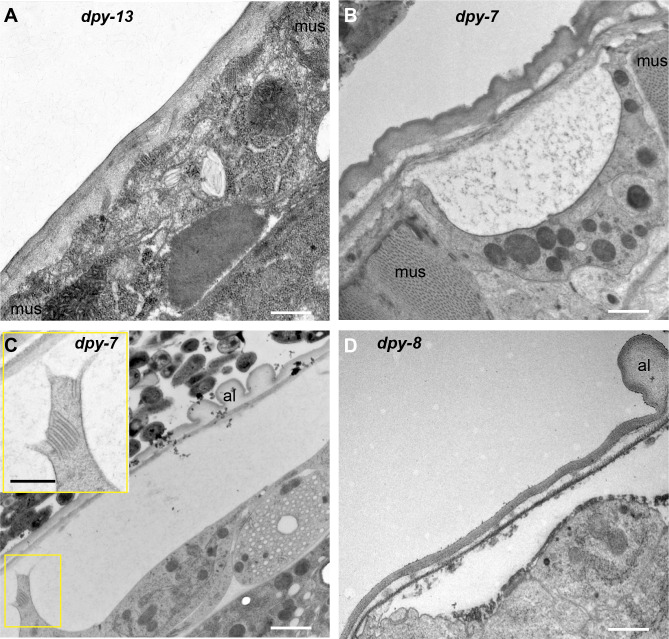
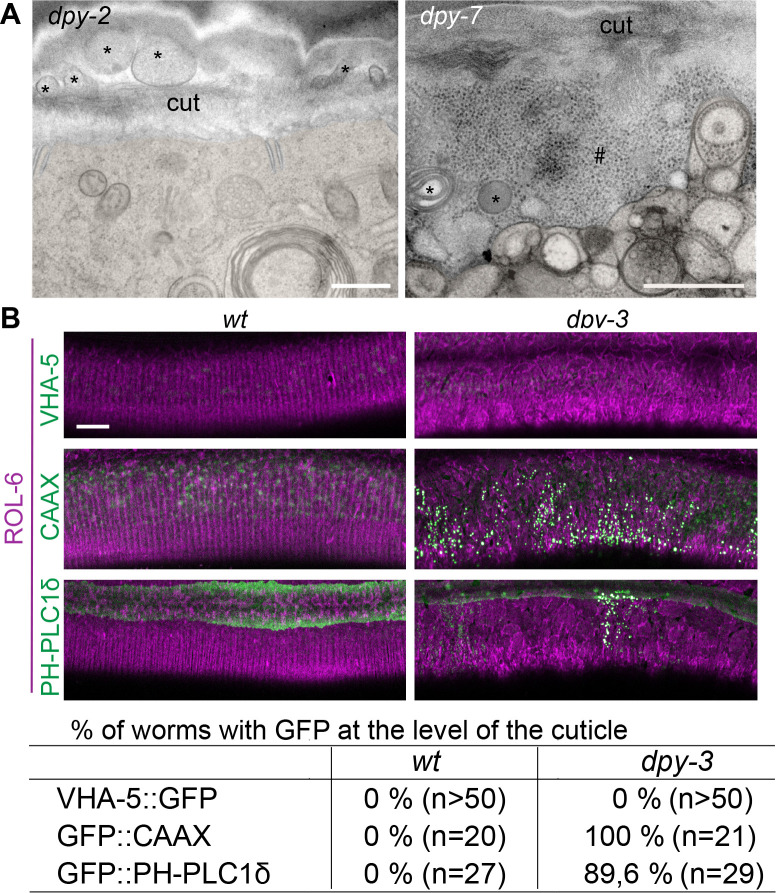
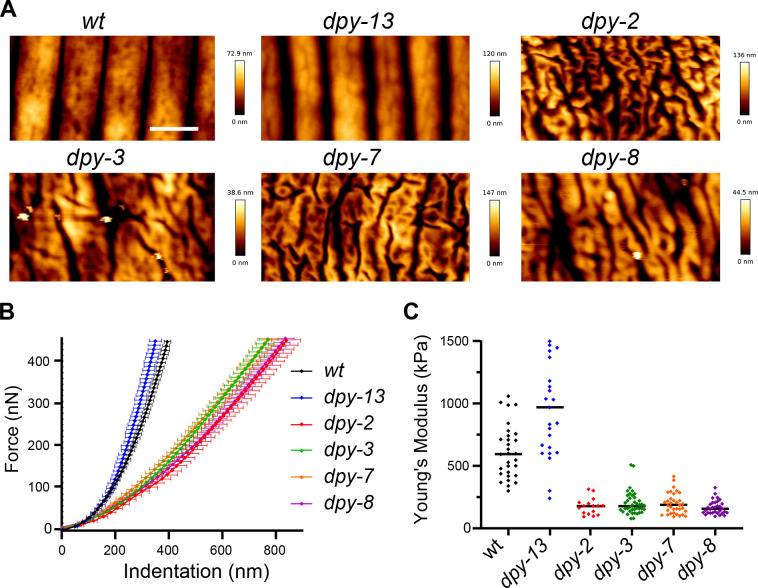
Apical extracellular matrices (aECMs) form a physical barrier to the environment. In Caenorhabditis elegans, the epidermal aECM, the cuticle, is composed mainly of different types of collagen, associated in circumferential ridges separated by furrows. Here, we show that in mutants lacking furrows, the normal intimate connection between the epidermis and the cuticle is lost, specifically at the lateral epidermis, where, in contrast to the dorsal and ventral epidermis, there are no hemidesmosomes. At the ultrastructural level, there is a profound alteration of structures that we term 'meisosomes,' in reference to eisosomes in yeast. We show that meisosomes are composed of stacked parallel folds of the epidermal plasma membrane, alternately filled with cuticle. We propose that just as hemidesmosomes connect the dorsal and ventral epidermis, above the muscles, to the cuticle, meisosomes connect the lateral epidermis to it. Moreover, furrow mutants present marked modifications of the biomechanical properties of their skin and exhibit a constitutive damage response in the epidermis. As meisosomes co-localise to macrodomains enriched in phosphatidylinositol (4,5) bisphosphate, they could conceivably act, like eisosomes, as signalling platforms, to relay tensile information from the aECM to the underlying epidermis, as part of an integrated stress response to damage.
Keywords: C. elegans; cell biology; cell–matrix interactions; damage; eisosomes; epidermis; extracellular matrix; host-defense system; signal transduction; skin.
© 2023, Aggad, Brouilly, Omi et al.
Conflict of interest statement
DA, NB, SO, CE, BD, CS, FR, CC, KP, DH, RP, NP No competing interests declared
Figures
Update of
References
-
- Bercher M, Wahl J, Vogel BE, Lu C, Hedgecock EM, Hall DH, Plenefisch JD. Mua-3, a gene required for mechanical tissue integrity in Caenorhabditis elegans, encodes a novel transmembrane protein of epithelial attachment complexes. The Journal of Cell Biology. 2001;154:415–426. doi: 10.1083/jcb.200103035. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
