

SOX17 Deficiency Mediates Pulmonary Hypertension: At the Crossroads of Sex, Metabolism, and Genetics
- PMID: 36913491
- PMCID: PMC10112457
- DOI: 10.1164/rccm.202203-0450OC
SOX17 Deficiency Mediates Pulmonary Hypertension: At the Crossroads of Sex, Metabolism, and Genetics
Abstract
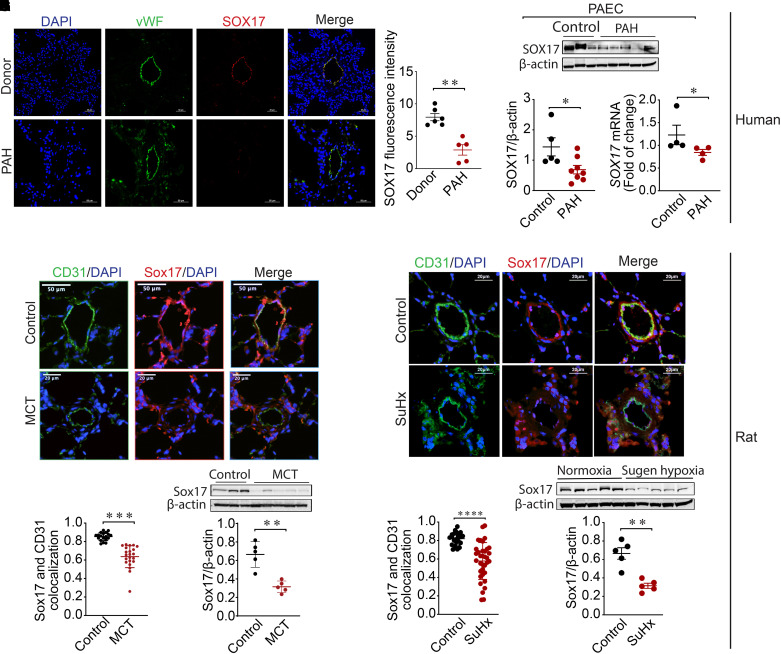
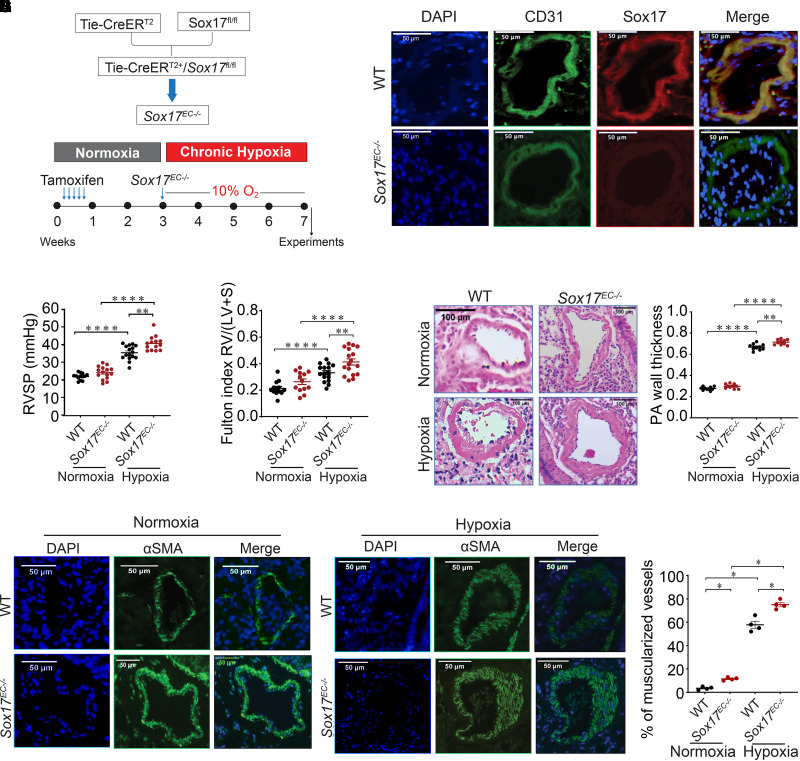
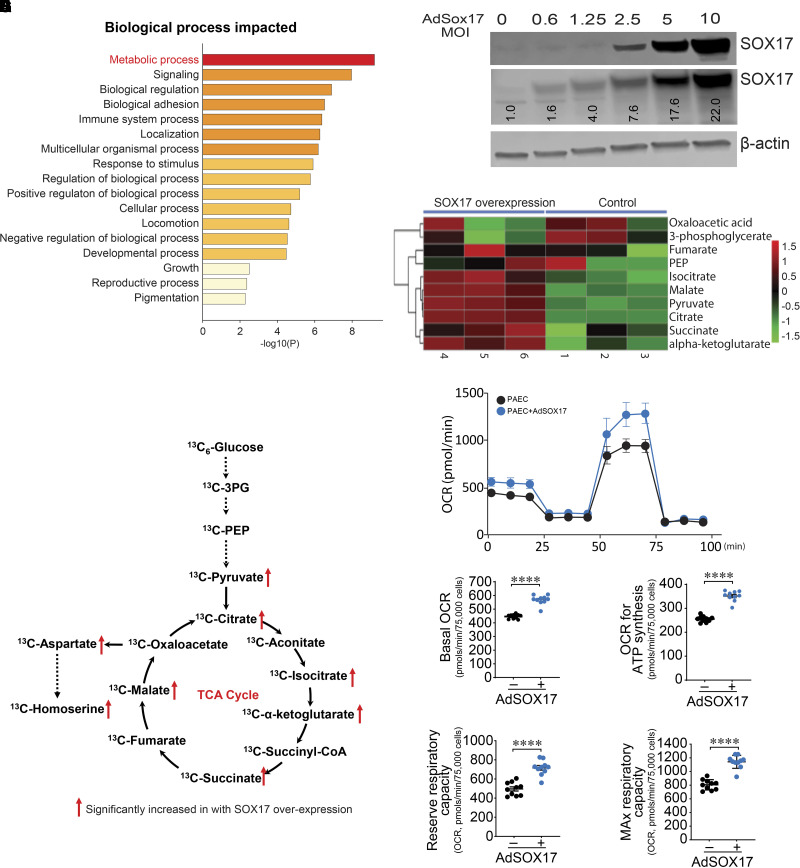
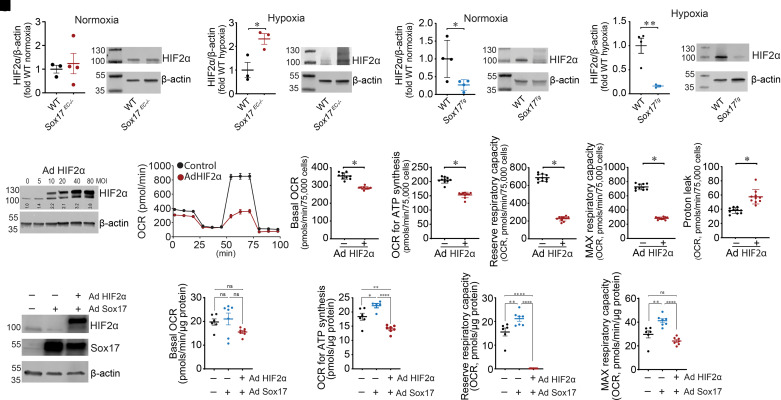
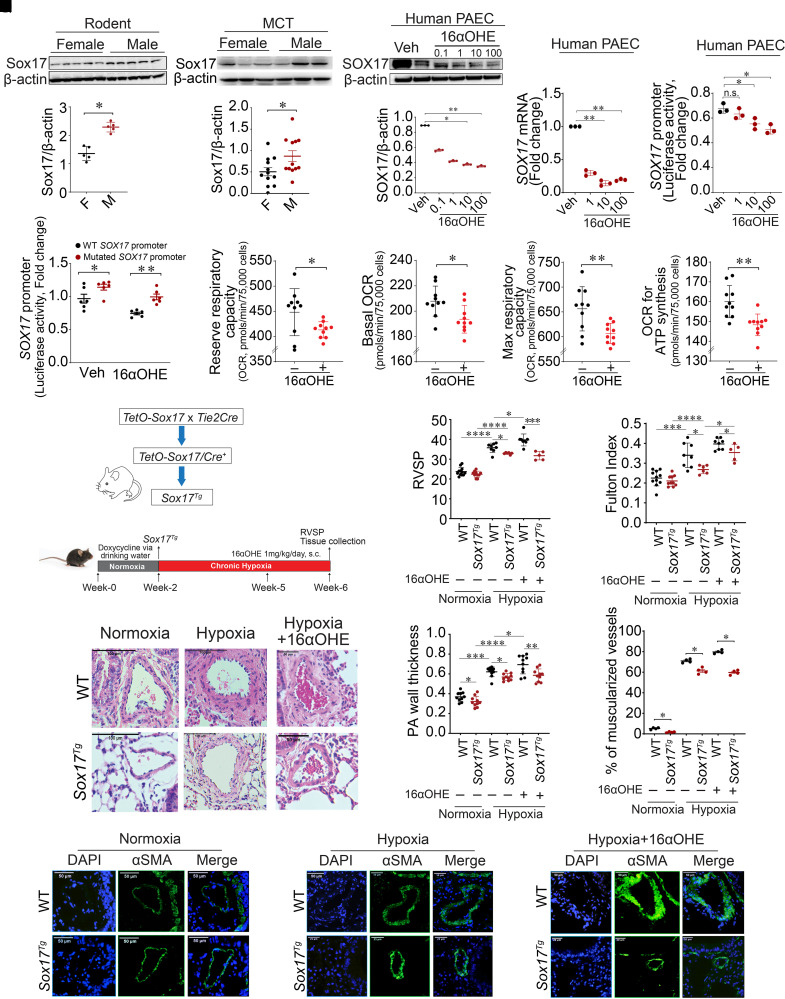
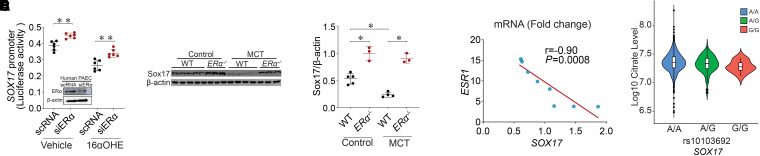
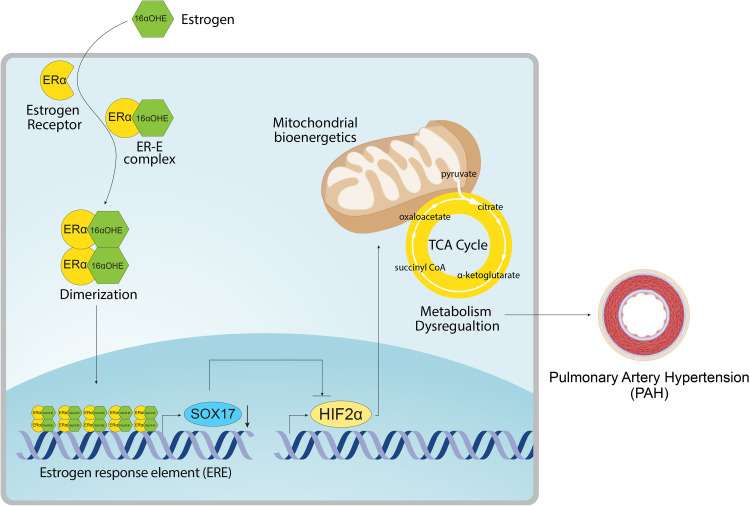
Rationale: Genetic studies suggest that SOX17 (SRY-related HMG-box 17) deficiency increases pulmonary arterial hypertension (PAH) risk. Objectives: On the basis of pathological roles of estrogen and HIF2α (hypoxia-inducible factor 2α) signaling in pulmonary artery endothelial cells (PAECs), we hypothesized that SOX17 is a target of estrogen signaling that promotes mitochondrial function and attenuates PAH development via HIF2α inhibition. Methods: We used metabolic (Seahorse) and promoter luciferase assays in PAECs together with the chronic hypoxia murine model to test the hypothesis. Measurements and Main Results: Sox17 expression was reduced in PAH tissues (rodent models and from patients). Chronic hypoxic pulmonary hypertension was exacerbated by mice with conditional Tie2-Sox17 (Sox17EC-/-) deletion and attenuated by transgenic Tie2-Sox17 overexpression (Sox17Tg). On the basis of untargeted proteomics, metabolism was the top pathway altered by SOX17 deficiency in PAECs. Mechanistically, we found that HIF2α concentrations were increased in the lungs of Sox17EC-/- and reduced in those from Sox17Tg mice. Increased SOX17 promoted oxidative phosphorylation and mitochondrial function in PAECs, which were partly attenuated by HIF2α overexpression. Rat lungs in males displayed higher Sox17 expression versus females, suggesting repression by estrogen signaling. Supporting 16α-hydroxyestrone (16αOHE; a pathologic estrogen metabolite)-mediated repression of SOX17 promoter activity, Sox17Tg mice attenuated 16αOHE-mediated exacerbations of chronic hypoxic pulmonary hypertension. Finally, in adjusted analyses in patients with PAH, we report novel associations between a SOX17 risk variant, rs10103692, and reduced plasma citrate concentrations (n = 1,326). Conclusions: Cumulatively, SOX17 promotes mitochondrial bioenergetics and attenuates PAH, in part, via inhibition of HIF2α. 16αOHE mediates PAH development via downregulation of SOX17, linking sexual dimorphism and SOX17 genetics in PAH.
Keywords: 16α-hydroxyestrone; SRY-related HMG-box 17; hypoxia-inducible factor 2α; metabolism; pulmonary arterial hypertension.
Figures
Comment in
-
SOX17 at the Intersection of Sex, Transcription, and Metabolism in Pulmonary Hypertension.Am J Respir Crit Care Med. 2023 Apr 15;207(8):971-972. doi: 10.1164/rccm.202302-0284ED. Am J Respir Crit Care Med. 2023. PMID: 36927296 Free PMC article. No abstract available.
References
-
- Rhodes CJ, Batai K, Bleda M, Haimel M, Southgate L, Germain M, et al. UK NIHR BioResource Rare Diseases Consortium; UK PAH Cohort Study Consortium; US PAH Biobank Consortium Genetic determinants of risk in pulmonary arterial hypertension: international genome-wide association studies and meta-analysis. Lancet Respir Med . 2019;7:227–238. - PMC - PubMed
-
- Wu Y, Wharton J, Walters R, Vasilaki E, Aman J, Zhao L, et al. The pathophysiological role of novel pulmonary arterial hypertension gene SOX17. Eur Respir J . 2021;58:2004172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL144727/HL/NHLBI NIH HHS/United States
- P01 HL134610/HL/NHLBI NIH HHS/United States
- R01 HL142212/HL/NHLBI NIH HHS/United States
- 1R01HL144727/HL/NHLBI NIH HHS/United States
- R01 HL158686/HL/NHLBI NIH HHS/United States
- R01 HL136603/GF/NIH HHS/United States
- I01 BX002042/BX/BLRD VA/United States
- R01 HL137282/HL/NHLBI NIH HHS/United States
- R01 HL156993/HL/NHLBI NIH HHS/United States
- P01 HL146369/HL/NHLBI NIH HHS/United States
- R01 HL160941/HL/NHLBI NIH HHS/United States
- R24 HL105333/HL/NHLBI NIH HHS/United States
- R01 ES030197/ES/NIEHS NIH HHS/United States
- R01 HL164791/HL/NHLBI NIH HHS/United States
- R01 HL060190/HL/NHLBI NIH HHS/United States
- R01 HL136603/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
